Özgün Adı: Hommel, B., Chapman, C. S., Cisek, P., Neyedli, H. F., Song, J. H., & Welsh, T. N. (2019). No one knows what attention is. Attention, perception & psychophysics, 81(7), 2288–2303. https://doi.org/10.3758/s13414-019-01846-w
Özet
Bu makalede, “dikkat”in (attention) tekil bir yapı (unitary construct) ve/veya sinirsel bir sistem olarak ele alınmasının yararlılığını sorguluyoruz. Dikkat aslında tek bir şekilde tanımlanmaktan ziyade pek çok anlamı barındırmaktadır. Bu kavramın hem açıklanması gereken olgular bütünü (explanandum) hem de bu olguları açıklayan süreçler bütünü (explanans) için kullanıldığını vurguluyoruz. Söz konusu argümanlarımızı örneklendirmek amacıyla, odağımızı görsel seçici dikkat (visual selective attention) olgusuna yöneltiyoruz. Evrim sürecinde, çok kanallı karmaşık bir duyu-motor (sensorimotor) sistemin doğal bir tasarım özelliği olarak seçiciliğin (selectivity) ortaya çıktığını; dikkatin de bu yapının ortaya çıkardığı yan ürünlerden biri olduğunu savunuyoruz. Dikkate yönelik geleneksel analitik yaklaşım yerine, başlangıçta dikkate özgü olmayan ama seçicilik süreçlerini açıklayabilen mekanizmalardan yola çıkan bütüncül bir yaklaşım öne sürüyoruz. Sonuç olarak, bilimsel ilerleme açısından en yararlı adımın, “dikkat” terimini belirli bir işlevsel ya da sinirsel sistemin etiketi olarak kullanmaktan vazgeçmek olduğunu düşünüyoruz. Bunun yerine davranışsal açıdan anlamlı seçim süreçlerine ve bunları hayata geçiren çok sayıdaki sisteme odaklanmayı öneriyoruz.
Anahtar Kelimeler: Dikkat (Attention) · Motor kontrol · Seçim (Selection) · Duyu-motor (Sensorimotor) · Karar verme (Decision making) · Filogenetik (Phylogenetic) · Niyet (Intention) · Evrim (Evolution) · Parietal korteks · Superior kollikulus
1. Giriş
“Herkes dikkatin ne olduğunu bilir” (James, 1890) ifadesi, William James’in en bilinen sözlerindendir ve şüphesiz insan dikkatine dair en ünlü ifadelerden biri olarak kabul edilir[1]. Ancak biz, bu ifadenin bilişsel araştırmalarda aşırıya kaçarak kullanılmasının ve popülerleşmesinin bilimsel ilerleme açısından olumsuz etkileri olduğunu; aslında kimsenin dikkatin ne olduğunu gerçekten bilmediğini savunmaktayız. Özellikle, dikkat kavramının bilişsel bilimlerde sıkça yanlış anlaşılan ve yanıltıcı bir şekilde kullanılan terimlerden biri olduğunu ileri sürmekteyiz.
Bu makalede şu görüşü savunuyoruz: Dikkat teriminin kullanımı bırakılmalı ve ilgili alandaki araştırmalar, belli bir göreve yönelik performansı sağlayan süreç ve mekanizma gruplarına odaklanarak yeniden yapılandırılmalıdır. Benzer görüşler daha önce de önerilmiş ve tartışılmıştır (bkz. Anderson, 2011; Di Lollo, 2018; Hommel & Colzato, 2015; Krauzlis, Bollimunta, Arcizet, & Wang, 2014; Mole, 2011). Çalışmamız, insanın duyu-motor (sensorimotor) bilgi işleme sistemlerinin birbiriyle bağlantılı ve bütünleşik (integrative) yapısına yeni bir odak getirerek önceki görüşleri hem doğrulamakta hem de geliştirmektedir. Duyu-bilişsel-motor süreçlerin (sensori-cognitive-motor processes) bütünleştirilmesine yapılan bu vurgu, bilişi (cognition) anlama konusunda geliştirilen sentetik yaklaşımlardan (Hommel & Colzato, 2015) ve davranışı anlamada filogenetik bir bilimsel perspektiften (Cisek, 2019) esinlenmektedir.
Bu makalede, öncelikle dikkat kavramının bugüne kadar nasıl kavramsallaştırıldığı ve çalışıldığı konusundaki temel sorunları tartışıyor ve özetliyoruz. Amacımız, bilişsel olguları incelerken sentetik bir yaklaşım benimsemektir. Bu yaklaşıma göre odak, dikkat kavramı altında incelenmiş süreçler ile mekanizmaların alt kümelerine yöneliktir. Yani dikkati tek bir bütünsel kavram olarak ele almak yerine, bu süreçleri ayrı ayrı anlamaya çalışmaktır. Durum analizimizi desteklemek amacıyla iki örnek sunuyoruz. İlkinde, dikkat ile niyet (intention) arasındaki kavramsal ayrımı ele alıyor ve mevcut tartışmanın verileri yeterince açıklayamadığını gösteriyoruz. İkinci örnekte ise seçim (selection) ve ödül geçmişi (reward history) araştırmalarını inceleyerek, geleneksel analitik yaklaşımların bu sorunu çözmede yetersiz kaldığını ortaya koyuyoruz. Analitik yaklaşıma alternatif olarak, insan beyninin filogenetik evrimini kısaca gözden geçiriyor (daha ayrıntılı inceleme için bkz. Cisek, 2019) ve seçici dikkatin (selective attention), hayvanların doğal ortamda karşılaştığı zorlukların bir sonucu olarak ortaya çıktığını gösteriyoruz. Son olarak, farklı ampirik gözlemleri tek bir “dikkat” kavramı altında toplamaya ve hepsini tek bir bütünsel sistem üzerinden açıklamaya yönelik geleneksel analitik yaklaşımın başarısız olduğu sonucuna varıyoruz. Bunun yerine, ekolojik açıdan anlamlı mekanizma ve süreçlerden başlayan ve bunlara odaklanan, ardından mümkün olduğunca çok sayıda olguyu (dikkate yönelik olan veya olmayan) açıklamayı hedefleyen daha sentetik bir yaklaşımın benimsenmesi gerektiğini savunuyoruz.
2. Dikkat Kavramı (The Concept of Attention)
Biz, dikkat kavramı ilgili sorunları gündeme getiren ilk araştırmacılar değiliz. Birçok yazar, dikkatin nesneleştirilme (reification) eğiliminin, ampirik sonuçların açıklanmasında tekrarlayan ve dolaylı bir mantık ürettiğini vurgulamıştır (Anderson, 2011; Di Lollo, 2018). Bir diğer yaygın eleştiri ise, genellikle dikkat olarak adlandırılan olguların altında birden fazla sürecin yattığı yönündedir (Di Lollo, 2018; Hommel & Colzato, 2015). Mole (2011), James’in ifadesinin, dikkatin esas işlevinin düşünme, algılama veya eylemde mi olup olmadığı tartışmalarının sürdüğü bir dönemde ortaya çıktığını vurgular. James’in çağdaşı F.H. Bradley, dikkat kavramına yönelik ilk eleştirilerden birini yapmış ve makalesine “Dikkatin Kendine Özgü Bir Faaliyeti Var Mıdır? (“Is There Any Special Activity of Attention?”)” başlığını vermiştir. Kısaca Bradley’nin görüşü, dikkat olarak etiketlenen çok sayıda olgu örneğinin mevcut olduğu, ancak bu olguların arkasındaki süreçlere yeterince önem verilmediği yönündeydi (Bradley, 1886).
Aradan 130 yılı aşkın bir süre geçmesine rağmen, dikkatin rolünü, dikkat kavramının kullanımını ve dikkatin ne olduğunu sorgulama ihtiyacı devam etmekte ve önem kazanarak artmaktadır (örn. Busemeyer, Gluth, Rieskamp, & Turner, 2019; Gottlieb, 2012). Biz de bu görüşleri yeniden teyit ediyor ve ayrıca, dikkati ayrı bölümlere ayırıp ardından tek bir dikkat sistemi (attentional system) arayışına girmeyi, insan davranışının kapsamlı bir biçimde anlaşılmasını engelleyen bir yaklaşım olarak görüyoruz. Çünkü bu yaklaşım, duyusal, bilişsel ve eylemsel süreçler arasındaki bütünleşik, paralel ve karşılıklı ilişkileri görmezden gelmektedir.
Temel görüşümüzü ve savlarımızı detaylandırmadan önce şunu belirtmek istiyoruz: Ele aldığımız kuramsal sorunlar özellikle dikkat kavramına yöneliktir, ancak yalnızca bu alanla sınırlı kalmamaktadır (Hommel, 2019a). Nitekim, ileride sunacağımız argümanlara oldukça benzer biçimde bellek kavramı da eleştirel sorgulamaya tabi tutulmuştur. İnsan belleği alanındaki uzun soluklu araştırmalar, belleğin performans boyutlarını ayrı ayrı temsil ettiği varsayılan çok sayıda bellek sistemi ortaya koymuştur. Bu sistemler kendi varlıklarını açıklayabilir gibi görünmekle birlikte altta yatan süreçlerin mekanistik olarak anlaşılmasına sağladıkları katkı oldukça sınırlı kalmıştır (Bechtel, 2008). Güncel çalışmalar, farklı bellek türlerinin birbirinden bağımsız ve özelleşmiş sistemlerin ürünü olmayabileceğini; bunun yerine normal işleyişteki bilişsel sistemlerin farklı sonuçları olarak ortaya çıktığını (Buckner & Schacter, 2004) ve insanlığın evrimsel sürecinin değişik aşamalarında geliştiğini ileri sürmektedir (Murray, Wise, & Graham, 2017). Benzer görüşler duygu (emotion) kavramı için de dile getirilmiş (Barrett, 2017; Hommel, 2019b) ve biliş de dahil olmak üzere diğer kavramlara da uyarlanabilmektedir (Cisek, 2019). Dikkat üzerine yoğunlaşmamızın sebebi, en azından bazı ilgili olguların duyusal, motor ve bilişsel etkileşimler açısından daha iyi anlaşılabileceği kanısında olmamızdır.
“Güncel çalışmalar, farklı bellek türlerinin birbirinden bağımsız ve özelleşmiş sistemlerin ürünü olmayabileceğini; bunun yerine normal işleyişteki bilişsel sistemlerin farklı sonuçları olarak ortaya çıktığını (Buckner & Schacter, 2004) ve insanlığın evrimsel sürecinin değişik aşamalarında geliştiğini ileri sürmektedir (Murray, Wise, & Graham, 2017).”
İnsan dikkati üzerine kuramsal yaklaşımlar en az üç temel sorunla karşılaşmaktadır. Birincisi, dikkat kavramı, dikkat olarak adlandırdığımız tüm süreçlere katkıda bulunan tek ve tutarlı bir bilişsel ya da sinirsel işlem dizisinin var olduğu yanılsamasını yaratabilmektedir– ki bu durum analiz düzeyine göre değişkenlik göstermektedir (örn. Kahneman, 1973). İkincisi, dikkat kavramı hem psikolojinin açıklamakla sorumlu olduğu kritik bir açıklanması gereken olgu (explanandum) hem de bu açıklamayı sağlaması beklenen açıklayıcı unsur (explanans) olarak yanlış yorumlanabilmektedir. Bu da açıklayıcı unsurun sahte açıklama (pseudo-explanation) karakteri kazanmasına neden olmaktadır. Üçüncüsü ise dikkat kavramının kararlar (decisions), niyetler (intentions), motivasyon (motivations), duygular (emotions), planlama (planning) ve hayata geçirme (execution) gibi diğer süreçlerden ayrılan özgün bir bilişsel veya sinirsel işlem grubunu (cognitive or neural operations) temsil ettiği düşünülmektedir. Sonraki bölümlerde bu üç noktanın detaylı tartışmasını yaparken göreceğimiz gibi, bu varsayımların hepsi yanlıştır.
İlk olarak, araştırmacıların tarihsel olarak dikkat kavramını kullanırken açıklamaya çalıştıkları olguların neler olduğunu ele alalım. Yaygın ve klasik görüşlere göre dikkat, bilgi işleme (process), belleğe kaydetme (store) ve hatırlama (retrieve information) kapasitemizi en verimli biçimde kullanmamızı sağlayan bilişsel ve sinirsel mekanizmaların bir bütünüdür. Bununla birlikte, giriş kitaplarına (örn. Eysenck & Keane, 2000) ve çevrimiçi kaynaklara bakıldığında, dikkatin çok farklı yetilerle ilişkilendirildiği görülmektedir: dış olayları daha derinlemesine içsel işlemden geçirmek amacıyla seçme becerisi (odaklanmış dikkat / focused attention); yanıltıcı ve/veya alakasız bilgileri görmezden gelme (seçici dikkat / selective attention); alakasız bilgileri işleme (istemsiz dikkat / involuntary attention); tek bir olaya ait bilgilerin aynı ya da farklı duyusal modaliteler arasında seçici olarak bütünleştirilmesi (özellik entegrasyonu / feature integration); belirli bir konumdaki olayların işlenmesine öncelik verilmesi (mekânsal dikkat / spatial attention); hedef bir olayı sistematik olarak arama (görsel arama / visual search); aynı anda birden çok iş yapma (bölünmüş dikkat / divided attention); göz hareketlerinin mekânsal parametrelerini kontrol etme (eyleme yönelik seçici dikkat / selective attention for action); bir hedefe diğerlerinden daha çok öncelik verme (hedef-merkezli dikkat / goal-centered attention); bir nesneye, bellek unsuruna veya bilinçli temsile diğerlerine kıyasla öncelik verme (nesne-merkezli dikkat / object-centered attention); bilgiyi daha sonra kullanılmak üzere pekiştirme ve olası bir olaya hazırlık olarak belli bir süre yoğunlaşma (sürdürülen dikkat / sustained attention). İlk bakışta, tüm bu olguların aynı işlevsel/sinirsel mekanizma grubu tarafından yürütüldüğünü varsaymak pek olası görünmemektedir (Allport, 1993). Davranışsal ve sinirsel alanlarda edinilen bulguların büyük bir kısmı da bu alt işlevlerin çoğunun birbirinden ayrıştırılabilir olduğunu doğrulamaktadır (örn. Fan, McCandliss, Sommer, Raz & Posner, 2002). Buna paralel olarak, bugüne dek “dikkat” başlığı altında toplanan bu farklı olguların hepsini tutarlı bir biçimde açıklayabilecek kapsamlı bir kuram henüz geliştirilmemiştir.
İkinci olarak, dikkat kavramı sıklıkla bilişsel işlemlemenin (cognitive processing) hem sorununu hem de çözümünü kapsamak için kullanılmaktadır. Diğer bir ifadeyle hem açıklanmak istenen olguyu hem de bu olguyu açıklamak üzere öne sürülen mekanizmayı tanımlar. Örneğin bu kavram, bir olayın temsiline (representation) diğer olayların temsillerine kıyasla daha fazla ağırlık kazandırarak karar verme ve eylem üzerinde daha güçlü bir etki yaratmasına yol açan istemli ve istemsiz faktörlerin sonuçlarını ifade etmek için kullanılmaktadır (örn. Yantis, 1998). Ancak aynı kavram, bu tür temsil farklılıklarının davranışı yönlendirmesinden doğan sonuçlarla başa çıkmayı sağlayan sistem, mekanizma veya yetiyi tanımlamak için de kullanılmaktadır (örn. Broadbent, 1958). Benzer şekilde, bazı araştırmacılar dikkati açıklanması gereken bilişsel darboğaz (the cognitive bottleneck), yani kritik kapasite sınırlaması olarak görürken (örn. Pashler & Johnston, 1998), diğerleri dikkati bu darboğazlarla baş etmeyi sağlayan bilişsel araç olarak değerlendirmektedir (örn. Bundesen, 1990). Bu kavramsal belirsizlikler, açıklanması gereken asıl sorunun ne olduğunu muğlaklaştırmıştır: Söz konusu olan, en iyi şekilde kullanmamız gereken bir bilişsel darboğaz mı, yoksa içinden seçilmesi gereken fazla miktarda bilgi midir? Dahası, dikkat kavramı sorunun kendisine mi yoksa çözümüne mi işaret etmektedir? Ortaya çıkan bu karışıklık, dikkat araştırmalarının ve kuramsal yaklaşımlarının, gözlenen olguların neden-sonuç ilişkilerini anlama temelli mekanistik anlayış yerine, döngüsel açıklamalara dayanması (örneğin, dikkat olgularının, dikkat sistemlerinin varlığı varsayılarak açıklanması) tehlikesini doğurmaktadır (Krauzlis ve al., 2014).
Üçüncü olarak, dikkat üzerine yapılan araştırmalar, psikolojik işleyişe yönelik yaygın analitik yaklaşımın etkisinde kalmış ve bu durumdan olumsuz etkilenmiştir (bu meseleye dair ayrıntılı bir tartışma için bkz. Hommel & Colzato, 2015). Analitik yaklaşım, her ne kadar dikkat alt işlevlerinin çeşitliliği göz önüne alındığında mümkün olmasa da kapsamlı bir tanım arayışındadır. Bu yaklaşıma göre varsayılan her bir dikkat alt işlevi (örneğin açık/overt ve örtük/covert, erken/early ve geç/late, odaklanmış/focused ve bölünmüş/divided, istemli/voluntary ve otomatik/automatic dikkat) ayrı işlevsel ve sinirsel süreçlerle ilişkilendirilmeli ve bu alanlarda yapılan araştırmalar süreçlerin kendisi yerine görevler ve alt işlevler üzerine yoğunlaşmalıdır. Bu yaklaşımın temel sorunu, dikkatin alt işlevleri ve hatta bilişe dair diğer kavramlar arasındaki ortaklıkları ele almamasıdır. Örneğin, dikkat, karar verme, niyet, duygu ve motivasyon kavramlarını farklı durum ve kuramsal bağlamlarda kullanıyor olmamız, bunların altında yatan işlevsel ve sinirsel süreçlerin gerçekten de farklı ve ayrışık olduğu anlamına gelmez. Nitekim, dikkatin temel süreçlerini diğerlerinden ayırmaya yönelik çabalar çoğunlukla ortak bir görüşe varılamadan sonuçsuz kalmıştır. Bir sonraki bölümde analitik yaklaşımın başarısızlığına örnek teşkil eden “dikkat” ve “niyet” kavramlarının birbirinden ayrıştırılmasına yönelik girişimleri irdeleyeceğiz.
3. Dikkat ve Niyet: Başarısız Bir Ayrım Girişimi
Çoğu araştırmacı, seçici dikkatin (selective attention) sinirsel temelinin posterior parietal korteksin olduğunda ve bu bölgenin “dikkat ağı” (attentional network) içinde kritik bir rol oynadığında hemfikirdir (Corbetta & Shulman, 2011; Posner & Dehaene, 1994; Ptak, 2012). Özellikle posterior parietal korteksteki her bir tekil nöron (individual neuron), dikkatin odaklandığı alanı yansıtmaktadır (Bisley & Goldberg, 2010; Robinson, Goldberg, & Stanton, 1978) ve bu bölgedeki hasar sıklıkla mekânsal ihmal (spatial neglect) olgusu ile ilişkilidir (Bartolomeo, 2007; Corbetta & Shulman, 2011). Buna karşın bazı araştırmalar, beynin bu bölgelerinin hareket kontrolü ile ilişkili süreçlerde de rol oynadığını göstermektedir (Mountcastle, Lynch, Georgopoulos, Sakata & Acuna, 1975; Snyder, Batista & Andersen, 1997). Özellikle posterior parietal korteksin bazı bölgelerinin, hareketin planlanması ve yönlendirilmesinde rol oynayan frontal lob ile güçlü ve karşılıklı bağlantıları mevcuttur (Johnson, Ferraina, Bianchi & Caminiti, 1996; Markov ve ark., 2014). Ayrıca tekil nöronların aktivitesi, aynı uyarıcılara yönelik gerçekleştirilen hareket türüne bağlı olarak düzenlenmektedir (Cui & Andersen, 2007; Snyder ve ark., 1997) ve posterior parietal korteksin inaktivasyonu, serbest seçim görevlerinde (free-choice tasks) sistematik önyargılara yol açarken (Christopoulos, Kagan & Andersen, 2018), görsel kanıtlara dayalı kararlarda ayni etkiyi yaratmamaktadır (Katz, Yates, Pillow & Huk, 2016).
Bu çelişkili bulgular, posterior parietal korteksin dikkati yönlendirmede mi yoksa niyeti yansıtmakta mı rol oynadığı konusundaki tartışmayı uzun yıllardır canlı tutmaktadır. Psikoloji bilimine hâkim analitik yaklaşımın tipik bir örneği olarak, dikkat ve niyet kavramları, birbirini dışlayan şekilde tanımlanmış ve bu iki kavramın deneysel olarak ayrıştırılabileceği varsayılmıştır. Bu tanımlamaya göre dikkat, duyusal bilgilerin bilişsel süreçlere erişimini kısıtlayan ve belirli bilgilerin bilinçli farkındalık kazanarak daha kapsamlı işleme tabi tutulmasını sağlayan süreçtir. Niyet ise, özgür irade olsun ya da olmasın, bilişsel süreçlerin ürünü olup belirli bir eylemi yapma yönündeki kararlılıktır. Oysa, dünyanın önde gelen sinir bilimcilerinin onlarca yıllık çabalarına rağmen, posterior parietal korteksin işlevinin net bir şekilde ayrıştırılması hâlâ mümkün olamamıştır. Culham ve Kanwisher (2001) bu durumu, mevcut parietal işlev hipotezlerinin gerçekte parietal lobların işlevsel örgütlenmesini yansıtmadığını ve kavramsal bir ilerleme olmadan daha iyi hipotezler test etmenin mümkün olmadığını ifade ederek özetlemiştir.
Bazı araştırmacılar, dikkat ve niyet arasındaki karmaşayı aşmak için daha bütünleştirici bir yaklaşım önermişlerdir; premotor teori (pre-motor theory) bu konuda öne çıkan bir modeldir. Premotor teoriye göre, dikkat kaymaları, göz hareketlerini yöneten (oculomotor) alt eşik (sub-threshold) sakkadik komutlarla tetiklenir ve tersine dikkatin mekânsal kaymaları hareket planlamasını başlatır (örn. Rizzolatti, Riggio, Dascola & Umilta, 1987). Bu fikre destek niteliğinde çok sayıda gözlem mevcuttur:
a) Dikkat ve göz hareketlerinin davranışsal olarak güçlü biçimde bağlantılı olduğunu gösteren davranışsal çalışmalar (örn. Deubel & Schneider, 1996; Kowler, Anderson, Dosher & Blaser, 1995; Sheliga, Riggio & Rizzolatti, 1995);
b) Görsel dikkat görevlerinde göz hareketi alanlarının aktivasyonunu gösteren fMRI çalışmaları (örn. Beauchamp, Petit, Ellmore, Ingeholm & Haxby, 2001; Corbetta ve ark., 1998; Nobre, Sebestyen & Miniussi, 2000) ve hareket iceren görevlerde retinotopik olarak tanımlanmış görsel kortekste hareket aktivasyonunun (dekodlama) gözlemlenmesi (Gallivan, Chapman, Gale, Flanagan & Culham, 2019);
c) Superior kollikulus (superior colliculus / SC), frontal göz alanı (frontal eye field / FEF) ve lateral intraparietal alanın (lateral intraparietal area / LIP) nöronlarının uyarılmasının dikkatin odak noktasını değiştirebileceğini gösteren çalışmalar (Cavanaugh & Wurtz, 2004; Cutrell & Marrocco, 2002; Moore & Fallah, 2001; Muller, Philiastides & Newsome, 2005);
d) Frontal korteks, parietal korteks veya orta beyin (midbrain) hasarına bağlı dikkat bozuklukları olan hastalar üzerinde yapılan nörolojik çalışmalar (örn. Husain & Kennard, 1996; Posner, Cohen & Rafal, 1982; Posner, Rafal, Choate & Vaughan, 1985; Sapir, Soroker, Berger & Henik, 1999).
Bu bağlamda özellikle önemli olan, yaklaşan bir sakkadın hedef konumunda algısal ayırma yetisinin iyileştiğini ortaya koyan davranışsal çalışmalardır (Deubel & Schneider, 1996; Gersch, Kowler & Dosher, 2004; Hoffman & Subramaniam, 1995; Peterson, Kramer & Irwin, 2004). Bu çalışmalar, sakkad başlamadan hemen önce, sakkad hedefindeki uyaranların öncelikli olarak işlendiğini göstermektedir; muhtemelen bunun nedeni dikkatin hedef konuma kaymış olmasıdır. Davranışsal çalışmaların ortaya koyduğu, yaklaşan bir sakkadın hedef konumunda algısal ayırma yetisini iyileştirdiği bulgusu bu bağlamda özellikle önem taşımaktadır (Deubel & Schneider, 1996; Gersch, Kowler & Dosher, 2004; Hoffman & Subramaniam, 1995; Peterson, Kramer & Irwin, 2004). Bu araştırmalar, sakkad hareketi başlamadan hemen önce, hedef noktadaki uyaranların öncelikli işlendiğini göstermektedir; bu durum dikkatsel odağın hedef noktaya önceden yönlendirilmesi ile açıklanabilir.
Başka araştırmalar, eylem ve dikkat arasındaki etkileşime dair çalışmaları el hareketlerini de kapsayacak şekilde genişletmiştir. Bu çalışmaların sonucunda, uzanma ile kavrama hareketlerinin planlanması ve gerçekleştirilmesinin bu hareketlerin yöneldiği nesnelerin işlenmesine öncelik verdiğini gösterilmiştir (örn. Pratt & Abrams, 1994; Rizzolati, Riggio & Sheliga, 1994; Tipper, Lortie & Baylis, 1992; ayrıca bkz. Wu, 2014). Gözler sabit kalsa bile, algısal ayırt etme hedeflenen konumda diğer konumlara göre daha etkili olmaktadır (Baldauf & Deubel, 2008; Baldauf, Wolf & Deubel, 2006; Deubel & Schneider, 2003; Deubel, Schneider & Paprotta, 1998; Khan, Song & McPeek, 2011). Gelecekteki eylemlere ilişkin niyetle bağlantılı “dikkatsel etki” (attentional impact) ya da öncelikli işleme (prioritized processing) sadece mekânsal önceliklendirme (spatial prioritization) ile sınırlı kalmamaktadır. Diğer çalışmalar, hareket etmeye hazırlanmanın veya hareket planlamanın, hedef nesnenin eylemle ilişkili özelliklerinin de algılanmasını kolaylaştırdığını göstermiştir. Örneğin, elimizle kavrama hareketine hazırlanmak boyut açısından farklı uyaranların (size oddballs) fark edilmesini, işaret etme hareketine hazırlanmak ise konum açısından farklı uyaranların (location oddballs) fark edilmesini kolaylaştırmaktadır (Fagioli, Hommel & Schubotz, 2007; ayrıca bkz. Craighero, Fadiga, Rizzolatti & Umiltà, 1999). Diğer bulgular da (Bekkering & Neggers, 2002; Moher, Anderson & Song, 2015; Tipper, Meegan & Howard, 2002; Weir ve ark., 2003; Welsh & Pratt, 2008; Welsh & Zbinden, 2009; ayrıca bkz. Gallivan, Barton, Chapman, Wolpert & Flanagan, 2015; Glazebrook, Welsh & Tremblay, 2016; Yoxon, Constable & Welsh, 2019) nesnelerin belirli özelliklerinin, o anda yapılacak eyleme özgü önemine bağlı olarak öncelik kazanabileceğini göstermektedir. Başka bir deyişle, belirli bir özellik (örneğin yönelim) bir durumda (örneğin kavrama eyleminde) öncelikli hale gelirken, başka bir durumda (örneğin işaret etme eyleminde) öncelikli olmamaktadır. Dolayısıyla, seçicilik yalnızca fiziksel uyaran özellikleri ya da eylem hedeflerinden kaynaklanmamaktadır. Seçicilik, bu faktörler arasındaki karşılıklı ve yinelenen etkileşimlerle şekillenmektedir. Bu bulgular, seçici dikkatin birden çok işlevsel ve sinirsel sistemi kapsadığını düşündürmektedir.
“Dolayısıyla, seçicilik yalnızca fiziksel uyaran özellikleri ya da eylem hedeflerinden kaynaklanmamaktadır. Seçicilik, bu faktörler arasındaki karşılıklı ve yinelenen etkileşimlerle şekillenmektedir. Bu bulgular, seçici dikkatin birden çok işlevsel ve sinirsel sistemi kapsadığını düşündürmektedir.”
Uyaran özellikleri ile eylem hedeflerinin etkileşimi, mekânların ve özelliklerin seçimi ile önceliklendirilmesinde rol oynamaktadır. Ancak bu seçimin yalnızca varsayımsal bir dikkat sistemi içinde gerçekleştiği açık değildir. Nitekim, potansiyel hedef konumlarda eyleme yönelik eş zamanlı aktif olan çoklu amaçlarla ve nihai hedef seçimiyle ilişkili sinirsel aktivite, eylem planlaması ve uygulanmasıyla daha yakın bağlantılı olan beyin bölgelerinde saptanmıştır. Bu yapılar dorsal premotor alan (dorsal premotor area), parietal uzanım bölgesi (parietal reach region) ve motor korteksi (motor cortex) içerir (Cisek ve Kalaska, 2005; Klaes, Westendorff, Chakrabarti ve Gail, 2011; Pesaran, Nelson ve Andersen, 2008; Scherberger ve Andersen, 2007; Song ve McPeek, 2010; Thura ve Cisek, 2014). Davranışsal açıdan, birden çok eşzamanlı tepki temsilinin bulunması ve hedef ile hedef-olmayan uyaran ve eylemler arasındaki dinamik seçim süreci, uzanma ve kavrama hareketlerinin mekânsal-zamansal özelliklerinde de ortaya çıkmaktadır. Dikkat temelli seçimin niyet oluşumundan önce tamamlanmış olmasıyla beklenebilecek etkili ve doğrudan hareketler yerine; el ve göz hareketlerinin yörüngeleri, hedef dışı uyaranların zamanlamasına ve öne çıkma derecesine bağlı olarak bu uyaranlara doğru veya onlardan uzağa kayma göstermektedir (örn. Chapman ve ark., 2010; Gallivan ve Chapman, 2014; Howard ve Tipper, 1997; Moher ve ark., 2015; Neyedli ve Welsh, 2012; Song ve Nakayama, 2006, 2008; Welsh, 2011; Welsh ve Elliott, 2004; Wispinski, Gallivan ve Chapman). Dolayısıyla, fiziksel olarak icra edilen eylemin özellikleri (characteristics of the physically executed action), hedef ve hedef dışı uyaranların “dikkatsel” durumunu doğrudan yansıtmaktadır. Bu bulgular, dikkat, seçilim (selection) ve niyetin ayrı ayrı kategoriler halinde ele alınamayacağını; bunların yoğun bağlantılar içeren paralel bir sistemde birlikte işleyen, süreklilik arz eden süreçler olduğunu göstermektedir.
Tüm bu sinirsel ve davranışsal araştırma bulgularını tutarlı bir çerçeveye oturtmak için daha fazla çalışma gerekli. Yine de bizim görüşümüze göre, dikkat ile niyet kavramlarının ayrımı burada ele aldığımız çeşitli bulguları açıklamada yetersiz kalıyor. Bu ayrım, bulguları anlamlı biçimde sınıflandırarak yeni kuramsal çalışmaları teşvik edecek bir kılavuz sağlamıyor ve giriş (input)/çıkış (output) süreçleri arasındaki çoklu etkileşimlerle de çelişiyor. Peki bu ve benzeri insan dikkatine dair kavramsal sorunlara (örneğin kontrollü (controlled) ve otomatik (automatic) süreçler veya kolaylaştırma (facilitation) ve baskılama (inhibition)) ne tür bir çözüm getirmek gerekir?
4. Başarısız Bir Analitik Çözüm: Seçim ve Ödül Geçmişi
Yukarıda belirtildiği gibi, psikolojik işleyişi çözümlemede hâkim olan analitik yaklaşım, bir kavramı eksiksiz biçimde tanımlayarak onu diğer kavramlardan kesin sınırlarla ayırmayı hedefler. Örneğin, “dikkat” kavramını anlamak için, bunun “niyet”, “karar verme” ya da “motivasyon” gibi kavramlardan nasıl farklı olduğu tartışılır. Ne var ki dikkat–niyet ayrımında görüldüğü üzere, bu strateji başarısızlıkla sonuçlanmıştır ve buna yönelik iki farklı alternatif önerilebilir. Birincisi, önceki tanımlama girişimlerinin yetersiz olduğu kabul edilip kavramlar arası örtüşmenin azaltılması hedeflenerek daha gelişmiş tanımlar önerilebilir. Örneğin dikkat kavramı yalnızca girdi (input) seçimine, niyet kavramını ise yalnızca çıktı (output) seçimine indirgenebilir. Dikkat kavramının bu denli daraltılması, artık bütünleştirme (integration), yönelim (orientation) ya da uyanıklık (vigilance) gibi diğer dikkate yönelik işlevlerle ilişkili görülmemesine, nihayetinde kavramdan vazgeçilmesini mecburi kılar. Aynı durum niyet kavramı için de geçerlidir. Olumlu yönden bakıldığında böyle bir yaklaşım, benzerlik göstermeyen süreçler ve yapılar arasında yapay ortaklıklar aranmasının önüne geçmektedir. Fakat olumsuz yanı, neden yalnızca bu işlevsel boyutların seçilip diğerlerinin dışlandığını açıklayacak hiçbir kuramsal temelin bulunmamasıdır. Bu nedenle, başlangıçta teknik bir tanım sorunu olarak beliren mesele, gerçekte gerekçesiz bir kuramsal önyargıya dönüşmektedir.
Başka bir alternatif olarak bazı araştırmacılar, ek bileşen ve faktörleri hesaba katan hibrit (hybrid) yaklaşımları çözüm olarak önermiştir. Buna örnek olarak 1970 ve 1980’lerde yoğun tartışmalara yol açan dikkat kaynakları kuramı (the resource theory of attention) (Kahneman, 1973; Navon, 1984) gösterilebilir. İlk yaklaşımlar, tüm zihinsel faaliyetlere paylaştırılması gereken tek bir kaynak varsayarak sade ve anlaşılır bir model veriyordu. Ne var ki yeni bulguların eklenmesiyle bunları birleştirme çabaları, giderek daha çok sayıda ve türde ayrı kaynak önerilmesine yol açtı. Sonuç itibariyle bu durum, sistematik öngörülerde bulunmayı olanaksız hale getirdi (Navon, 1984). Bu yaklaşım günümüzde büyük ölçüde terk edilmiştir; yalnızca ego tükenmesi (ego-depletion) alanında benzer tartışmaların sürdüğü görülmektedir (Friese, Loschelder, Gieseler, Frankenbach ve Inzlicht, baskıda). Hibrit modellerin başarısızlığının asıl nedeni ise, bir araya getirilen farklı faktörlerin tutarlı ve bütünleşik bir kuramsal çerçeveye dâhil edilememesidir.
Seçici dikkat konusunda da benzer bir yönelim görülmektedir. Awh, Belopolsky ve Theeuwes (2012), kişi tarafından yönlendirilen ya da içsel dikkat (endogenous attention) ile mevcut eylem ve hedeflerle ilgili olmayan uyarıcıların öncelikli işlenmesini gösteren dış kaynaklı ya da dışsal dikkat (exogenous attention) arasındaki farkı ele almışlardır. Aynı araştırmacılar bu iki türü arasındaki geleneksel ayrımla artık uyuşmayan bulgular arasında bağlantı kurma girişiminde bulunmuşlardır. İçsel-dışsal dikkat ayrımı, tıpkı dikkat-niyet ayrımı gibi, kavramları daha iyi tanımlama amacıyla yapılmış; fakat giderek artan kanıtlar, bu kategorilerin yetersizliğini açığa çıkarmıştır. Özetle, içsel dikkat bazen fazlasıyla otomatik, dışsal dikkat ise hedefe ve çevresel uyaranların özelliklerine bağımlı olabileceği için bu ayrım, uygulamada sıklıkla işlevsiz kalmaktadır (Awh ve ark., 2012; Folk, Remington ve Johnston, 1992; Hommel ve Wiers, 2017). Awh ve arkadaşları (2012) bu sorunu çözmek amacıyla üçüncü bir değişken olarak seçim geçmişi (selection history) eklenmesini önermektedir. Bu yaklaşıma göre hedeflerin (içsel dikkatin temel belirleyicisi), belirginliğin/salıens’in (salience) (dışsal dikkatin temel belirleyicisi) ve seçim geçmişinin (önceki seçimler ile bu seçimlerin sonucu olan ödüllerle ilişkili, söz konusu içsel-dışsal ikiliğine uygun olmayan bir faktör) tümü seçiciliğe (selectivity) katkı sağlamakta, çıktılar (output) ise bütünleştirici bir öncelik haritasına (integrative priority map) yönlendirilmektedir. Bu yaklaşım birçok mevcut bulguyu açıklayabilir görünse de bizim kanaatimizce asıl sorunu çözmemekte, kavramsal bir bütünlükten ziyade geçici bir “yama” (patch) işlevi görmektedir. Sonuçta bu yaklaşım uygun çözümlerin aranmasını engellemekte veya bu alandaki araştırmaları yanlış yönlendirmektedir. Bizim görüşümüze göre, radikal bir çözüm olarak bu kavramın bütünüyle tasfiyesi ve yeniden inşası gereklidir.
İlk bakışta, insan merkezi sinir sisteminde öncelikli işlemeyi belirleyen bilgilerin toplanıp bütünleştirildiği özel yapı veya yapılardan söz etmekte herhangi bir sorun yok gibi görünmektedir. Bu bağlamda öne çıkan yapılardan biri, görsel alan için öncelik haritası oluşturduğu düşünülen superior kollikulus’tur (superior colliculus) (Fecteau ve Munoz, 2006). Superior kollikulus’un yalnızca okülomotor kontrolle sınırlı olmadığı; göz, baş, vücut ve elin yönelim hareketlerinde de rol oynadığı gösterilmiştir (Gandhi ve Katnani, 2011; Stuphorn, Hoffmann ve Miller, 1999). Bu veriler, davranışsal bakımdan önemli olan uyaranlar için seçiciliğin tüm hareketli bedene yayıldığı görüşü ile tutarlılık gösterir. Song ve arkadaşlarının bulguları, özellikle el ile uzanma görevlerinde superior kollikulus’un hedef seçimindeki nedensel rolünü göstererek bu görüşü pekiştirmektedir (Song, Rafal, ve McPeek, 2011; Song ve McPeek, 2015). Böylece bu yapının genel amaçlı bir hedef seçme/yönelim (selection/orientation) sistemi içinde işlev gördüğü fikrini desteklemişlerdir (Nummela ve Krauzlis, 2010; Song, 2011). Öte yandan, superior kollikulus’un dikkati yönlendiren bilgilerin bütünleştiği tek harita olduğunu düşünmek gerekmediği gibi, mevcut tüm bilgilerin burada toplandığını varsaymaya da gerek yoktur. Bir sonraki kısımlarda da tartışacağımız üzere, insan beyninde çok sayıda seçicilik kaynağı vardır ve nihayetinde bütünleştirmeyi gerçekleştiren yapı beynin tamamıdır. Bu bütünleştirme süreci zaten açıklanması gereken bir olgudur (explanandum). Dolayısıyla, sadece bu işlevi yapmak için tasarlanmış tek bir haritanın var olduğunu söylemek, psikolojik bir olguyu açıklamak için o olguyu bir nevi yaratmakla görevli özel bir sistem olduğunu varsaymak anlamına gelir.
“Dolayısıyla, sadece bu işlevi yapmak için tasarlanmış tek bir haritanın var olduğunu söylemek, psikolojik bir olguyu açıklamak için o olguyu bir nevi yaratmakla görevli özel bir sistem olduğunu varsaymak anlamına gelir.”
Bu daha genel üst-kuramsal (meta-theoretical) sorun dışında bütünleştirmenin nasıl gerçekleştiğini açıklamadan sadece varlığını varsayan modellere yeni bir faktör eklemek, araştırmaları anlamlı biçimde yönlendirmeyecektir. Örneğin Awh ve arkadaşlarının (2012) yaklaşımında, seçim geçmişi hem hedef odaklı içsel hem de belirginlik (salience) odaklı dışsal seçicilikle büyük ölçüde örtüşmektedir. Belirli eylem türlerini planlamak ve uygulamak, nesnelerin bazı özelliklerinin işlenmesini sistematik olarak kolaylaştırır (örn. elimizle kavramada boyut ve yönelim, bir nesneye uzanmada konum: Bekkering ve Neggers, 2002; Craighero ve ark., 1999; Fagioli ve ark., 2007). Bu kolaylaşma, genetik olarak belirlenmiş olmaktan ziyade, yaşam boyu kavramaya dair edinilen deneyim ve öğrenme sürecinin bir sonucudur (Hommel, 2010). Gerçekten de kavrama eyleminde şekil ve yönelime odaklanmak, renge odaklanmaktan daha işlevseldir; çünkü başarılı bir kavramayı belirleyen temel özellikler bunlardır. Dolayısıyla, kavrama sırasında şekil ve yönelime renk karşısında bir öncelik vermek işlevsel bir uyum (functional adaptation) olacaktır. Bununla birlikte seçim geçmişinin, hedeflerin içsel dikkati yönlendirme biçimini şekillendirdiği görülmektedir. Aynı şekilde, görsel özelliklerin belirginliğinin çocukluğun ilk yıllarında ciddi biçimde değişmesi (örn. Suchman ve Trabasso, 1966), seçim geçmişinin belirginliği (salience) de etkileyebileceğini düşündürmektedir.
Her ne kadar tüm bu görüşler, Awh ve arkadaşlarının (2012) önerdiği seçim geçmişinin üçüncü bir faktör olarak dikkate alınmasını desteklese de aslında bu üç faktörün birbirinden bağımsız değil, hem ampirik hem de kavramsal düzeyde güçlü biçimde iç içe geçtiğini göstermektedir. Açıklamaya çalıştığımız üzere, bu tür kavramsal örtüşme sorunlarının daha fazla tanım üretmekle çözülebilmesi pek olası değildir. Bunun yerine, yalnızca bütünleştirmenin gerçekleştiğini varsayan değil, bu bütünleştirmenin isleyişini de açıklayan bir kurama ihtiyaç vardır.
Seçim geçmişini ek bir faktör olarak dahil etmenin daha çok soru işaretini beraberinde getirmesinin bir başka nedeni de kavramın kendisinin belirsiz olmasıdır. Özellikle içsel ve dışsal dikkatin dışında da pek çok faktörle örtüşmesi, kavramsal ayrışmayı zorlaştırmaktadır. Bu faktörlerden biri, seçim geçmişiyle iç içe olan ödül geçmişidir (reward history). Daha önce ödüllendirilmiş uyaranların öncelikli işlem gördüğü tartışmasız bir bulgudur (Anderson, Laurent, ve Yantis, 2011; Anderson ve Yantis, 2012). Bu da ödül geçmişinin belirginliği (salience) şekillendirmede önemli olduğunu göstermektedir. Normalde, bir organizmanın ödül alabilmesi için uyaranların önceden seçilmiş olması gerekir; bu da seçim geçmişi ve ödül geçmişi kavramlarının iç içe geçmesine yol açmaktadır. Awh ve arkadaşları (2012) da bu konuya dikkat çekmekte ve hem önceki ödülleri hem de önceki uyaran seçimlerini “seçim geçmişi”nin (selection history) örnekleri olarak değerlendirmektedir. Bununla birlikte, seçim geçmişi ve ödül geçmişi aynı şey olamaz; zira önceki seçimlerin tümü ödülle sonuçlanmış değildir. Dahası, ödül geçmişinin bu ortak kategoriye dahil edilmesi, teorik bütünleştirmenin öne sürdüğü kadar açık değildir; çünkü ödül geçmişinin kendisi büyük olasılıkla birbirinden ayırt edilebilir pek çok faktörden oluşmaktadır.
Örneğin, ödüllendirici uyaranlara karşı tercihimiz değişkenlik gösterir. Yüksek ödül alma şansı varsa bu tercih daha güçlüdür; başarısızlık maliyeti düşükse daha zayıftır (Neyedli ve Welsh, 2015b). Bu bulgu, katılımcıların ödüllendirici olarak algıladıkları şeyin çok boyutlu olduğunu düşündürmektedir. Aslında yalnızca ödülün büyüklüğü değil, aynı zamanda ödülü alma olasılığı da önem taşımaktadır. Ayrıca, bir dizi ulaşma-karar deneyinde (reach-decision experiments) çok sayıda önyargılayıcı (biased) faktör gözlemlenmiştir. Bunlardan bazıları ödül değeri ve olasılığı (Chapman, Gallivan ve Enns, 2015a), karar vermeye yönelik en iyi seçenek (Wispinski, Truong, Handy ve Chapman, 2017), mevcut birikmiş servet düzeyi (Neyedli ve Welsh, 2015a), hedeflerin nasıl algılandığından ziyade miktarı (Milne, 2013) ve olası seçeneklerinin nasıl sunulduğu (Chapman, 2014) olarak sayılabilir.
Buradaki temel sorun, Awh ve arkadaşlarının (2012) çözmeye çalıştığı soruna benzemektedir, o da incelenen kavramın tanımının fazla dar tutulmasından kaynaklanmaktadır. Daha önce de açıkladığımız gibi içsel ve dışsal kontrol ayrımı yeterli olmadığından Awh ve arkadaşları (2012) seçim geçmişini eklemişlerdir. Ne var ki seçim geçmişi dahi kendi içinde seçim (selection) ve ödül geçmişi (reward history) olarak ayrıştırılabilir ve hatta ödül geçmişi de farklı alt birimler çerçevesinde düşünülebilir. Dolayısıyla, ödülün doğası ve etkisi, çok sayıda bağlamsal değişkene bağlı olarak farklı şekillerde ortaya çıkmaktadır. Bu nedenle, tekil bir yapı önerisi açıklayıcı olmaktan uzaktır.
Eşit büyüklükteki kayıp ve kazanç arasındaki simetriyi sorgulayan çalışmalar, kavramsal tabloyu daha da karmaşıklaştırmaktadır. Örneğin, Kahneman ve Tversky’nin (1979) çalışmalarıyla ünlü hale gelen kayıp aversiyonu (loss aversion), eşit büyüklükteki kayıpların kazançlardan daha ağır bastığını öne sürmektedir. İlginç bir şekilde, hızlı kararlar alınacağı zaman bu etki tersine dönmekte (Chapman, 2015b), seçim davranışında yine asimetriler gözleniyor olsa da kayıp aversiyonunun tersi gözlenmektedir. Bir başka ifadeyle, bu tür durumlarda katılımcılar orantısız bir şekilde kazanç sağlayan seçeneklere yönelirken, kayıpla ilişkili tercihlerin caydırıcılığı zayıflamaktadır.
Bu sonuçlar bizi başka bir karmaşık duruma götürüyor: farklı önyargılar (biases) muhtemelen farklı zaman aralıklarında ortaya çıkmaktadır. Örneğin, iyi bir seçeneği tercih etmek ile kötü bir seçenekten kaçınma kararlarındaki asimetriyi inceleyen bir çalışmada, önyargıların işleme hızları farklılık göstermektedir. Bireyler, kazanç sağlayan hedeflere yönelimde kayıplardan kaçınmaya kıyasla daha hızlı, 100 ms daha erken tepki verirler (Chapman, 2015b). Bu durum, olumsuz değere dayalı görsel-motor seçimlerin optimal yapılabilmesi için ödül olasılığına dayalı farklılıklara kıyasla daha fazla işlem süresine ihtiyaç duyulduğunu öne süren çalışmalarla da tutarlıdır (Neyedli ve Welsh, 2015b).
Tüm bu bulgular birlikte ele alındığında, seçim geçmişinin ödül geçmişiyle yakından iç içe geçtiğini ve bu kavramların hiçbirinin yeterince iyi anlaşılmadığı görülmektedir. Bu nedenle seçim geçmişini, halihazırda tam olarak anlaşılmamış olan dışsal ve içsel dikkat kavramlarına eklemek, seçici dikkatin mekanizmalarını anlamaya fazla katkı sağlamayacaktır. Bizim önerimiz yeni bir teori inşa etmekten ziyade, teorik çerçevenin değişmesidir. Karmaşık olguları dikkatle tanımlayıp daha basit bileşenlerine ayırarak açıklamayı hedefleyen analitik yaklaşım yerine, basit mekanizmaların ve işlevsel süreçlerin etkileşiminden karmaşık davranışsal olguların nasıl ortaya çıktığını irdeleyen sentetik bir bakış açısı benimsemek gerektiğine inanıyoruz.
5. Sentetik Bir Yaklaşım
Sentetik bir yaklaşım, yalnızca biyolojik gerçeklikle uyumlu sinirsel ve işlevsel süreçleri bütünleştirdiğinde anlam taşır. Ne var ki, bu süreçlerin neler olduğunu belirlemek zordur. Yapacağımız sentezi biyolojik zeminde tutmanın etkili yollarından biri ise evrimi bir referans noktası olarak kullanmaktan geçmektedir. Bu doğrultuda “filogenetik arıtma” (phylogenetic refinement) adı verilen bir yöntem kullanılabilir. “Filogenetik arıtma” prosedürü, evrimsel çizgide sinirsel ve davranışsal süreçlerin nasıl geliştiğine ilişkin teorilerin, her evrimsel aşamada ortaya çıkan değişim ve uyumlarla uyumlu olacak şekilde giderek ayrıntılandırılmasını içerir (bkz. Cisek, 2019). Bu nedenle, burada dikkat kavramından bir süreliğine uzaklaşıp Homo sapiens’e, yani insana giden soy hattında ilgili sinirsel devrelerin nasıl evrimleştiğine dair kısa bir tarihsel inceleme yapıyoruz. Her ne kadar evrimde belirli bir değişikliğin neden ortaya çıktığını bilmek çoğu zaman güç olsa da hangi değişikliklerin hangi sırayla gerçekleştiğini ortaya koymak, karşılaştırmalı ve gelişimsel verilerin zenginliğine dayanır. Bu da sinirsel devrelerin ve davranışsal yetilerin nasıl birlikte evrimleştiğine ilişkin güçlü ve test edilebilir hipotezler üretmeyi mümkün kılar.
Primat soy hattındaki mekânsal etkileşim, uzun ve karmaşık bir evrimsel süreçle şekillenmiştir (Şekil 1). Yaklaşık 500 milyon yıl önce Kambriyen döneminde (Cambrian epoch) görsel rehberli yönelim davranışlarının ortaya çıkması önemli bir adım olmuştur. İlkel kordalı atalarımız, tek bir fotosensitif yama (photosensitive patch) aracılığıyla orta beyne (tektum) projeksiyon yapan ve omuriliğe bağlanarak kaçış tepkilerini tetikleyen basit bir görsel kaçış devresine sahipti (Lacalli, 1996, 2018). Omurgalı soy hattında, fotosensitif yama başın iki yanına, iki farklı göz yaması olacak şekilde ayrıldı (Butler, 2000). Göz yamaları, karşı taraftaki tektuma projeksiyon yapıyor ve tektum ise aynı taraf omuriliğine bağlantı sağlıyordu. Bu sinirsel devre, atalarımızın yaklaşan avcılardan uzaklaşmasını sağlayan reflekse dayalı kaçış hareketlerini mümkün kıldı (Şekil 2a). Göz yamaları genişledikçe, kubbe şeklini alarak lensi oluşturdu ve böylece dış uyaranların topografik haritalamasını sağlayan iki boyutlu bir retina ortaya çıktı (Lamb, 2013). Tektum da paralel olarak genişledi; yüzeysel katmanlarında üç boyutlu topografik bir harita oluşurken, derin katmanlarda ise projeksiyon gradyanları (gradients of downstream projections) oluşturdu. Sonuç olarak, belirli konumlardaki tehdit uyaranlarına karşı yönelimli kaçış tepkilerini temsil eden bir “eylem haritası” (action map) oluştu.
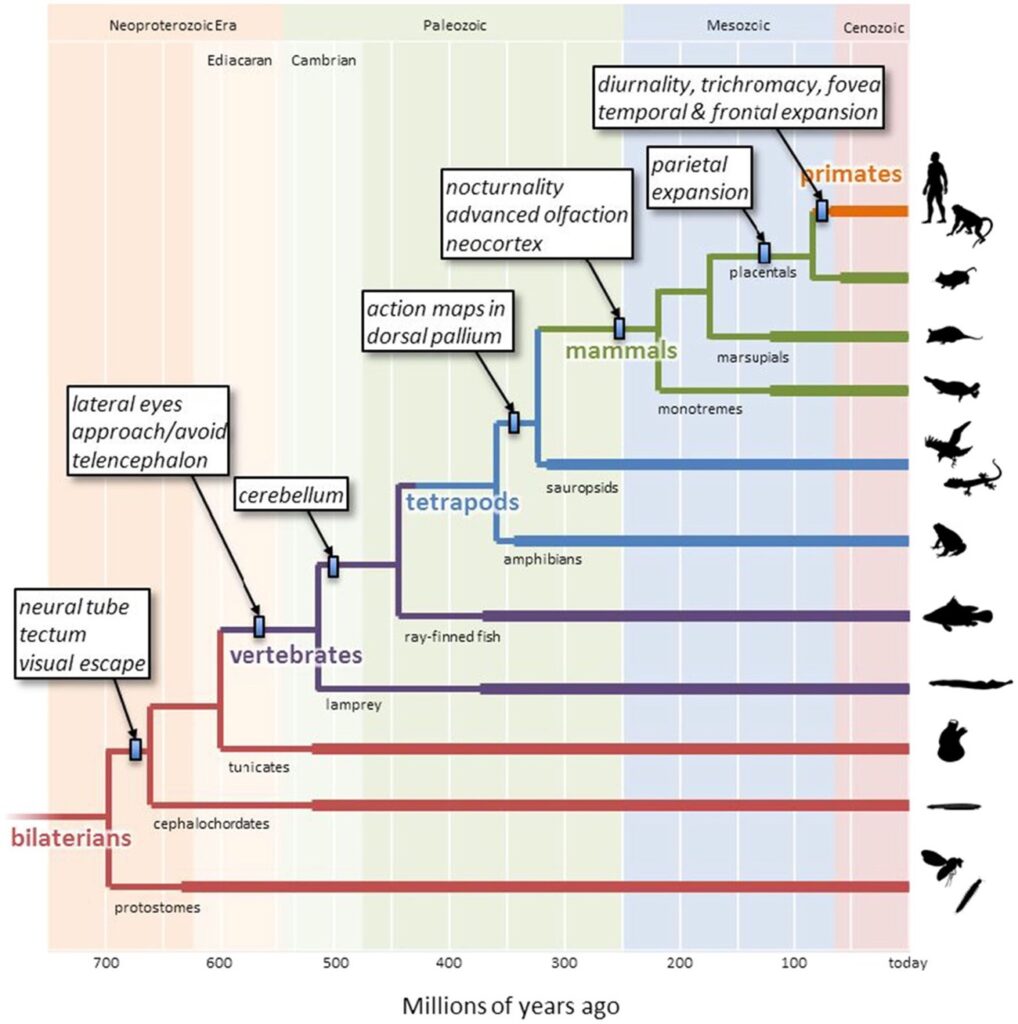 Şekil 1. İnsanlara giden soy hattını özel olarak vurgulayan, bilateral simetrili hayvanların sadeleştirilmiş bir filogenetik ağacı. Dallanma noktaları, farklı soy hatları arasındaki bazı ayrışmaları temsil eder ve zamanlamalar moleküler saat analizlerine dayanarak tahmin edilmiştir (Erwin ve ark., 2011). Kalın çizgiler, ilgili fosil verilerinin varlığını gösterir (paleobiodb.org). Küçük dikdörtgenler, kutularda belirtilen yeniliklerin tahmini en geç zamanını gösterir. Açıklık için birçok dallanma noktası ve soy hattı çıkarılmıştır. Sağ taraftaki siluetler phylopic.org’dan alınmıştır.
Şekil 1. İnsanlara giden soy hattını özel olarak vurgulayan, bilateral simetrili hayvanların sadeleştirilmiş bir filogenetik ağacı. Dallanma noktaları, farklı soy hatları arasındaki bazı ayrışmaları temsil eder ve zamanlamalar moleküler saat analizlerine dayanarak tahmin edilmiştir (Erwin ve ark., 2011). Kalın çizgiler, ilgili fosil verilerinin varlığını gösterir (paleobiodb.org). Küçük dikdörtgenler, kutularda belirtilen yeniliklerin tahmini en geç zamanını gösterir. Açıklık için birçok dallanma noktası ve soy hattı çıkarılmıştır. Sağ taraftaki siluetler phylopic.org’dan alınmıştır.
At nalı balığı (lamprey) çenesiz bir balık olup, atalarımızdan yaklaşık 550 milyon yıl önce ayrılmıştır. Bu balıklar üzerinde yapılan mikrostimulasyon (microstimulation) çalışmaları, tektumlarında yönelimli kaçış tepkilerinin organize bir haritasının varlığını ortaya koymaktadır (Saitoh, Menard ve Grillner, 2007). Bu çalışmalar ayrıca tektumun ön (rostral) bölgesinde yer alan başka bir eylem haritasının da varlığını göstermektedir. Bu harita, hayvanın önündeki mekâna duyarlıdır ve çoğunlukla karşı taraf omuriliğe projeksiyon yapar; böylece yönelim ve yaklaşma davranışlarını yönlendirmektedir (Jones, Grillner, ve Robertson, 2009; Kardamakis, Saitoh, ve Grillner, 2015). Tektumun bu bölgesi, dikkat ve seçilim açısından kritik öneme sahiptir. Kaçış devresinde (avoidance circuit), birden fazla uyaran birden fazla kaçış hareketini tetikleyebilir ve bu hareketlerin ortalaması alınarak uygun bir kaçış davranışı oluşturabilir (Şekil 2b). Öte yandan, yaklaşma devresinde (approach circuit) ortalama almak işe yaramaz çünkü iki uyaranın ortalama yanıtı, hayvanın her ikisini de gözden kaçırmasına neden olur (Şekil 2c). Bu nedenle, yaklaşma devresinde bir eylem diğerini tamamen bastıracak şekilde seçim yapılmalıdır. Bu seçim, yanal inhibisyon mekanizmaları aracılığıyla “kazanan hepsini alır” tarzı bir dinamikle (“winner-take-all” dynamics) gerçekleştirilebilir (Grossberg, 1973; Mysore ve Knudsen, 2011; Wang, 2002).
“Öte yandan, yaklaşma devresinde (approach circuit) ortalama almak işe yaramaz çünkü iki uyaranın ortalama yanıtı, hayvanın her ikisini de gözden kaçırmasına neden olur.”
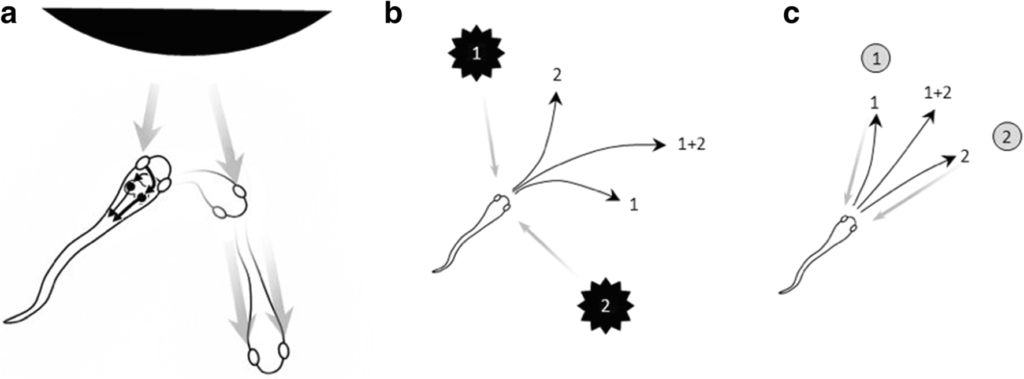 Şekil 2. Erken dönem bir omurgalıda kaçınma ve yaklaşma devreleri.
Şekil 2. Erken dönem bir omurgalıda kaçınma ve yaklaşma devreleri.
(a) Kaçınma devresinde, yanal gözlerden gelen görsel bilgi karşı taraftaki tektuma ulaşır ve buradan aynı taraftaki orta beyin hareket bölgelerine iletilir. Dolayısıyla, bir uyaran sol göze düştüğünde, hareket sağa doğru yönelir ve bu yönelim, uyarım dengelenene kadar sürer; böylece organizmanın bedeni uyarandan uzağa çevrilmiş olur.
(b) İki tehdit edici uyarandan (siyah yıldızlar) uzaklaşma yönlerinin (numaralı oklar) mekânsal ortalaması, etkili bir kaçış tepkisi üretir.
(c) Yaklaşma davranışlarında ise mekânsal ortalama işlevsizdir; bu nedenle “kazanan hepsini alır” (winner-take-all) prensibi gereklidir.
B ve C panelleri, Cisek (2019)’dan izin alınarak yeniden kullanılmıştır.
Peki, tüm bunlar dikkat ile nasıl ilişkilidir? Girişte alıntıladığımız ünlü ifadeden sadece birkaç cümle sonra James, dikkatin “bir şeylerden geri çekilmeyi (withdrawal) ve böylece diğer şeylerle etkili bir şekilde ilgilenmeyi içerdiğini” yazmıştır. Bazı uyaranlardan geri çekilme ve başka bir uyaranla etkileşime girme süreci, gerçekten de ön (rostral) tektumun yaklaşma devresi içerisinde gerçekleşir. Bu basit devreler, insanlardaki daha yüksek düzey bilişten oldukça uzak görünse de aslında “seçici dikkat” olarak adlandırılan mekanizmaları kontrol eden sistemlerin öncülleridir. Tektum, insan beynindeki superior kollikulus ile homolog bir yapıdır. Superior kollikulus’un, hem göz ve baş hareketleriyle bakış yönelimini düzenlemede hem de bakış sabitken örtük dikkati (covert attention) kontrol etmede kritik rol oynadığı bilinmektedir (Basso ve May, 2017). Soy hattımızın Kambriyen’in erken dönemlerinde minogillerden (lamprey) ayrılmasından bu yana merkezi sinir sistemi ve çevresel koşullar önemli ölçüde değişmiş olsa da tektumun yaklaşma ve kaçınma devreleri hem balıklarda (Herrero, Rodriguez, Salas, ve Torres, 1998) hem de memelilerde (Comoli ve ark., 2012) korunmuştur.
Zamanla atalarımız denizlerden ayrılmış ve bunların bir kısmı olan amniyotlar, tamamen karasal bir yaşam biçimine uyum sağlamıştır. Bu uyum süreci, koku, görsel ve dokunsal bilgileri bütünleştiren ve zamanla serebral korteksi (cerebral cortex) oluşturacak olan telensefalik palliumun (telencephalic pallium) genişlemesi ve katmanlaşması ile eş zamanlı gerçekleşmiştir (Aboitiz ve Montiel, 2015; Striedter, 2005). Tüm memelilerde neokorteks, uzamsal olarak topografik özellik gösteren dorsomedial bölüm (dorsomedial sector) ve topografik olmayan ventrolateral bölüm (ventrolateral sector) olmak üzere iki tabakadan oluşur (Finlay ve Uchiyama, 2015). Primatlarda dorsomedial bölüm, medial ve dorsolateral prefrontal korteks, singulat bölgeler, tüm premotor, motor, duyusal-motor ve parietal korteksin yanı sıra retrosplenial korteksi kapsar. Ventrolateral bölüm ise lateral prefrontal korteksin bazı bölümlerini, orbitofrontal korteksi ve tüm limbik korteks ile temporal lobu içerir. Dikkat açısından özellikle önemli olan bölge dorsomedial neokortekstir. Bu yapı, türlere özgü eylem sınıflarını desteklemek üzere organize olmuş bir dizi fronto-parietal devre içerir (Graziano, 2016; Kaas ve Stepniewska, 2016).
Yaklaşık 300 milyon yıl önce yaşayan erken memelilerde bu sistem büyük olasılıkla ilkel bir yapıdaydı ve yalnızca, lokomosyonla/hareketle (locomotion) ilişkili medial devreler ile baş ve ağız hareketleriyle ilişkili lateral devrelerden oluşuyordu (Kaas, 2017). Bu devrelerin her biri, kendine özgü bir biçimde duyusal bilgiyi işliyor ve belli bir eylem türüne özgü şekilde uzmanlaşıyordu (örneğin, lokomosyon/hareket için bacakların çevresindeki alan, beslenme için burnun çevresindeki alan). Ayrıca her devre, kendi isleyişine uygun belirli efektör gruplarına (a specific set of relevant effectors) sinyal gönderiyordu. Bir bakıma, her bir devre yaklaşma ve kaçınmada rol oynayan daha eski tektal sistemlere benzer bir “eylem haritası” (action map) niteliğindeydi. Ne var ki bu haritalar, memelilerin ekolojik koşullarında mevcut olan çok daha geniş bir görev yelpazesini yönlendiriyordu.
Memelilerin davranış çeşitliliği arttıkça dorsomedial neokortekste de genişleme oldu; bu da eyleme-özgü duyusal mekân haritalarının farklılaşmasını beraberinde getirdi. Primatlarda parietal korteksin genişlemesi özellikle çarpıcı olmuş ve farklı eylem türlerinin gereksinimlerine özgü çeşitli özgün mekân temsilleri ortaya çıkmıştır (Andersen, Snyder, Bradley ve Xing, 1997; Stein, 1992) (bkz. Şekil 3). Örneğin, görsel girdilerle yönlendirilen uzanma davranışları medial intraparietal korteks tarafından kodlanır (Cui ve Andersen, 2007; Kalaska ve Crammond, 1995). Bu bölge, erişilebilir mesafedeki hedefleri bakış yönü ve elin pozisyonuna göre kodlayarak temsil eder (Buneo, Jarvis, Batista ve Andersen, 2002; Gallivan, Cavina-Pratesi ve Culham, 2009). Ayrıca uzanmayı yöneten frontal devrelerle, özellikle dorsal premotor korteksle de bağlantılar kurar (Johnson et al., 1996; Wise, Boussaoud, Johnson ve Caminiti, 1997). Kavrama eyleminin kontrolünde anterior intraparietal alan öne çıkar (Baumann, Fluet, ve Scherberger, 2009). Bu alan, nesne şekline duyarlıdır ve ventral premotor korteks gibi kavramaya özgü frontal bölgelerle bağlantılıdır (Nakamura, 2001; Rizzolatti ve Luppino, 2001). Bakış yönünün kontrolü ise lateral intraparietal alan tarafından sağlanır (Snyder, Batista, & Andersen, 2000). Bu alan ise görsel mekânı göz retinasına göre (retinotopic frame) temsil eder (Colby ve Duhamel, 1996; Snyder, Grieve, Brotchie ve Andersen, 1998) ve frontal göz alanları ile superior kollikulus gibi bakış kontrolünden sorumlu yapılarla etkileşim halindedir (Paré ve Wurtz, 2001). Böylece, kökeni Kambriyen çağındaki tektal yönelim devrelerine dayanan eski bir evrimsel mekanizma yeniden devreye girer.
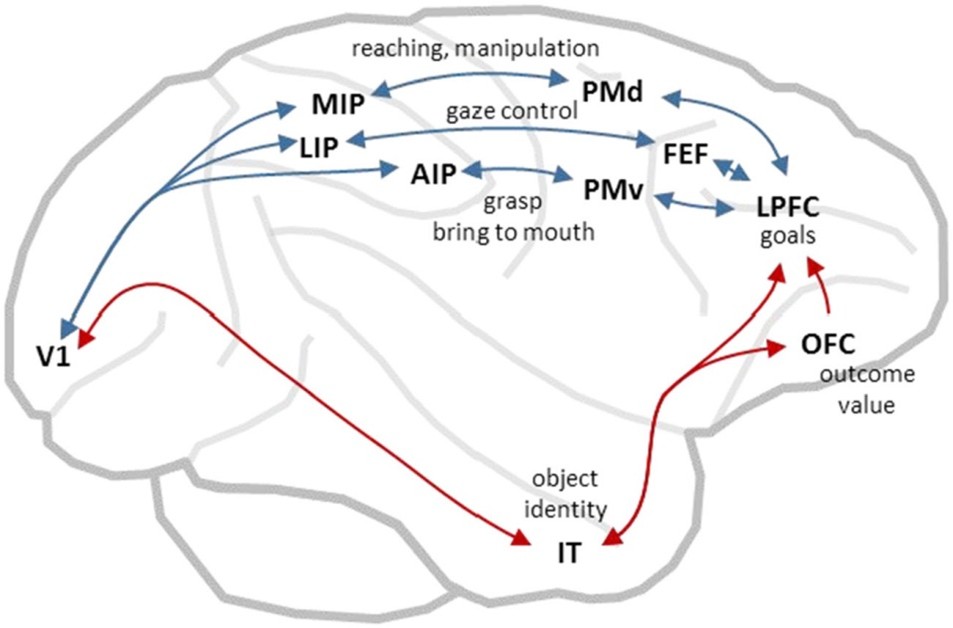 Şekil 3. Primat beyin korteksi, dorsomedial bölgelerde (mavi oklar) birbirine paralel bir dizi sensörimotor akış içerir. Bu bölgelerin her biri, uzamsal temsil biçimlerine dayalı olarak, belirli bir eylem türüne aracılık eder. Ayrıca ventrolateral bölgelerde (kırmızı oklar) hesaplanan nesne kimliği ve sonuç değeri bilgilerini kullanarak mevcut davranışsal bağlamda en uygun eylemleri seçer. Kısaltmalar: AIP (anterior intraparietal alan), FEF (frontal göz alanları), IT (inferotemporal korteks), LIP (lateral intraparietal alan), LPFC (lateral prefrontal korteks), MIP (medial intraparietal alan), OFC (orbitofrontal korteks), PMd (dorsal premotor korteks), PMv (ventral premotor korteks), V1 (birincil görsel korteks).
Şekil 3. Primat beyin korteksi, dorsomedial bölgelerde (mavi oklar) birbirine paralel bir dizi sensörimotor akış içerir. Bu bölgelerin her biri, uzamsal temsil biçimlerine dayalı olarak, belirli bir eylem türüne aracılık eder. Ayrıca ventrolateral bölgelerde (kırmızı oklar) hesaplanan nesne kimliği ve sonuç değeri bilgilerini kullanarak mevcut davranışsal bağlamda en uygun eylemleri seçer. Kısaltmalar: AIP (anterior intraparietal alan), FEF (frontal göz alanları), IT (inferotemporal korteks), LIP (lateral intraparietal alan), LPFC (lateral prefrontal korteks), MIP (medial intraparietal alan), OFC (orbitofrontal korteks), PMd (dorsal premotor korteks), PMv (ventral premotor korteks), V1 (birincil görsel korteks).
Çoğu durumda, farklı fronto-parietal eylem akışları birbirleriyle rekabet eder. Örneğin, bir canlının bir ağacın köklerini kazmak ya da bunun yerine başka bir ağaca doğru yürümek arasında “hepsi ya da hiç” (all-or-none decision) şeklinde bir karar vermesi gerekebilir. Ancak diğer bazı durumlarda, farklı fronto-parietal diziler eş güdümlü/koordine hale gelir. Örneğin, bir baş/burun yönelimi sistemi bir hedefi işaret ettiğinde, o hedef artık kazma veya ısırma gibi diğer davranışlar için kullanılabilir hale gelir. Bu kullanılabilirlik özelliği, ağaç tırmanıcı böcek yiyicilerden evrimleşen, merkezde yüksek çözünürlüklü bir fovea (görme çukuru) bulunan büyük gözler geliştiren ve meyve yeme alışkanlığı edinen primatlar için özellikle önem kazanmıştır. Bu tür hayvanlarda bakış yönelimini kontrol eden sistem, diğer birçok görsel rehberli davranış için bir yönetici rol (executive role) üstlenir. Bakış için bir hedef seçmek, neye uzanılacağını veya tırmanmak için hangi dalın kavranacağını seçmenin bir parçası haline gelir. Bu sistem, günümüzde “seçici dikkat”e atfedilen rolün büyük bir kısmını yerine getirir. Nitekim hem açık/overt hem de örtük/covert seçici dikkatin, bakış yönelimi sistemiyle yakından ilişkili olduğu ve aynı sinirsel yapıları içerdiği uzun süredir bilinmektedir (Corbetta ve ark., 1998; Rizzolatti ve ark., 1987). Bu yapılar arasında arka parietal korteks, frontal göz alanları ve superior kollikulus yer alır.
Bu kısa primat evreni yolculuğu bizi yine seçici dikkate ve posterior parietal kortekse getirmiş oldu, ancak bu kez her ikisine de farklı bir perspektiften bakıyoruz. Sorun, posterior parietal korteksin “dikkat” adı verilen bir süreçte mi yoksa “niyet” adı verilen başka bir süreçte mi rol oynadığı değildir. Artık esas soru, bu yapının, hayvanların hedeflerine ulaşmalarını ve olumsuz sonuçlardan kaçınmalarını sağlamak için çevreleriyle olan etkileşimleri seçmelerine ve kontrol etmelerine olanak tanıyan daha geniş bir sistem içinde nasıl bir yer edindiğidir. Filogenetik açıdan bakıldığında, primatların posterior parietal korteksin, nesnelere yönelik görsel etkileşimleri yöneten topografik dorsomedial neokortikal sistemin bir parçasıdır (Cisek, 2007). Bu sistem, paralel duyu-motor diziler (parallel sensorimotor streams) halinde organize olmuştur. Her bir dizi, hayvanın davranışsal dağarcığındaki belirli bir eylem türüne katkıda bulunur ve bunların etkinliği, bazal gangliyonlar ve diğer yapılardan gelen seçici teşvik (selective invigoration), enerji sağlama (energization) veya motivasyon (drive) mekanizmalarıyla düzenlenir (Cisek ve Thura, 2018; Grillner, Robertson ve Stephenson-Jones, 2013). Bu fronto-parietal eylem dizilerinin her birinde hedef seçimi, “kazanan hepsini alır” (winner-take-all) dinamiğinde gerçekleşir. Bu süreç, her bir eylem türü için özgül olan, kendine has bir mekânsal referans çerçevesi içinde (örneğin, göz hareketleri için retinotopik, yani göz retinasına göre) meydana gelir. Bu dizilerden biri, göz ve baş hareketleri yoluyla bakışı yönlendirmekle ilgilidir ve diğer birçok dizinin, foveadan sağlanan yüksek çözünürlüklü görsel bilgiye dayanması nedeniyle bir yönetici rol (executive role) üstlenmiş gibi görünmektedir. Bir hayvanın (insanlar da bu kategoriye dahil) laboratuvar koşullarında karmaşık doğal davranışın yalnızca tek bir bileşenini gerçekleştirme talimatı aldığını varsayalım. Posterior parietal kortekste gözlemlenen etkinlik, deneysel olarak manipüle edilen belirli görev değişkenlerine bağlı olarak, araştırmacının “dikkat”, “niyet” veya “karar verme” olarak tanımladığı şeyle ilişkili gibi görünür. Ancak, bu ilişkilerden (correlations) yola çıkarak beyinde bağımsız bir “dikkat sistemi” (veya benzer şekilde bir “niyet sistemi” ya da “karar sistemi”) var olduğu sonucu çıkarılamaz. Buradaki kilit çıkarım şudur: Posterior parietal korteks, bir “dikkat sistemi”nin parçası değildir. Tam tersine seçici dikkat, posterior parietal korteksin asıl işlevi olan hedefe yönelik eylemleri yönetme sürecinin doğal bir ürünüdür.
“Posterior parietal korteks, bir “dikkat sistemi”nin parçası değildir. Tam tersine seçici dikkat, posterior parietal korteksin asıl işlevi olan hedefe yönelik eylemleri yönetme sürecinin doğal bir ürünüdür.”
6. Özet ve Sonuçlar
Sonuç olarak, seçiciliğin evrim boyunca, hedefe yönelik eylemlerin verimli bir şekilde yürütülmesini sağlamak amacıyla ortaya çıkan bir tasarım özelliği olarak geliştiğini söyleyebiliriz. İnsana uzanan organizma soyunun eylem dağarcığı genişledikçe, bu tür bir seçicilik gerekli hale gelmiştir. Bu durum, seçiciliğin altta yatan sayısız süreçten doğan bir ortaya çıkan özellik olduğu anlamına gelir. İnsanların (ve seçici dikkat gösteren diğer türlerin) evrimleşme biçiminin basit bir gerçeğidir; seçici dikkat, “seçici niyet” ve “seçici karar verme” ile birlikte, bu sürecin birçok yan ürününden yalnızca biridir. Bu yazıda ağırlıklı olarak superior kollikulus ve parietal korteksteki seçim mekanizmalarını vurguladık, ancak benzer argümanlar diğer beyin bölgelerindeki diğer seçim mekanizmaları için de geçerlidir. Örneğin, Krauzlis ve arkadaşları (2014), bazı “dikkat” olgularının, bazal gangliyonların değer temelli seçim mekanizmalarının bir ürünü olabileceğini öne sürmektedir. Eğer seçicilik, davranış dağarcığının sayıca ve karmaşıklık bakımından (örneğin kaçınma, yaklaşma, sakkad, yeme, uzanma, kavrama ve alet kullanma) artmasıyla evrimleşen bir tasarım özelliği ise, davranışsal seçimi tek bir özelleşmiş sinirsel veya işlevsel sistemle açıklamaya çalışmak yetersiz kalmaktadır. Bizce, dikkat araştırmalarında seçici dikkatin gerçek kökenleri ve süreçleri hakkında bu kadar çok uzun soluklu ve oldukça verimsiz tartışmanın yaşanmasının nedeni de budur. Bu tartışmalar genellikle ikili (binary) bir nitelik taşır; çünkü tek bir neden veya merkezi bir sistemin varlığı varsayımıyla başlar. Bu tek neden, ek araştırma bulgularıyla giderek daha fazla sorgulanır, genellikle zıt bir başka neden ortaya atılır ve böylece tartışmalar devam eder. Bizim öngörümüz, bu tartışmaların hiçbir zaman sona ermeyeceği yönündedir. Bunun nedeni, seçicilik evrim boyunca pek çok faktörün etkileşiminden doğan bir özellik olduğu için, tüm tarafların savunucularının bir şekilde ve bazı durumlarda haklı olabileceğidir.
Yine de bu tartışmaların alanımızı daha ileri taşımadığına ve “dikkat olgularının” nasıl oluştuğuna dair anlayışımızı gerçekten derinleştirmediğine kesinlikle inanıyoruz. Analitik yaklaşımı sürdürmek ve tüm bu bulguları tek bir kavram olan “dikkat” ile açıklamaya çalışmak (yani tek bir açıklayıcı değişken olarak kullanmak) sorun yaratacaktır. Dikkat kavramı kaçınılmaz olarak o kadar çok farklı bulguya yayılır ki, sonunda ampirik bir ağırlığı kalmayacak kadar belirsiz hale gelir. Araştırmacıların bunları çok sayıda açıklanması gereken olgu (explananda) olarak ele almaları doğru olsa da yalnızca tek bir açıklayıcı ilke aramak ya da bunu zorunlu kılmak yanlış olur. Bunun yerine, mümkün olduğunca, ilgili temel mekanizma ve süreçleri belirlemeye ve her birini ayrı ayrı açıklamaya çalışmak gerekir.
Bilimsel analitik yaklaşımda, ortaya atılmış kavramların esiri olma riski vardır. Birçok araştırmacı, “dikkat”, “niyet” ve “karar verme” gibi kavramları günlük dilden alır ve bu dilsel kategorilerin bir şekilde beyindeki tanımlanabilir mekanizmalara veya işlevlere karşılık gelmesini bekler. Oysa gerçek sinirsel işlevlere bakıldığında net bir ayrım yoktur, yalnızca sonunda seçiciliği ortaya çıkaran bir dizi etkileşen süreç vardır. Bu süreçler, özel bir sisteme ait oldukları için değil, insan beyni ve vücudu bu şekilde evrimleştiği ve seçiciliğin etkin davranışı sağlamak için gerekli bir özellik olduğu için etkileşir. Dahası, bir bireyin yaşamı boyunca (uzak ve yakın geçmişte) yaptığı her şey, seçimi yaratır, pekiştirir ve şekillendirir. Örneğin sola dönmek, sağdaki uyaranları görmezden gelmemize neden olur; bir elmayı seçmek, diğerlerini gözden kaçırmamıza yol açar; bir kelime söylemek, başka bir kelime söylememizi engeller. Her bir seçim, pozitif kazançlardan negatif kayıplara kadar farklı şekillerde sonuçlanır. Dolayısıyla seçim ve ödül, yaşamlarımızın ve yaşam tarzımızın ayrılmaz parçalarıdır (Allport, 1987).
“Dolayısıyla seçim ve ödül, yaşamlarımızın ve yaşam tarzımızın ayrılmaz parçalarıdır (Allport, 1987).”
Seçici davranışın ortaya çıkması için, birbiriyle bağlantılı çok sayıda süreç, çeşitli bilgi kaynaklarını bütünleştirir. Bu süreçlerin farklı zaman çerçevelerinde gerçekleşmesi önemli bir zorluğu meydana getirir (örn. Chapman ve ark., 2015b; Welsh, Neyedli, ve Tremblay, 2013). Bu nedenle, bir laboratuvar ortamında, bu süreçler yalnızca seçim işlemi sırasında yalnızca tek bir noktada veya anlık olarak gözlemlendiğinde, yapılan gözlem “dikkat,” “niyet” veya “karar verme ve ödül” olarak yorumlanabilir. Burada önerilen sentetik yaklaşım, ödül ve seçim geçmişinin iç içe geçmiş konular olduğunu açık bir şekilde ortaya koyarken, muhtemelen hedef odaklı davranışa katkıda bulunan çoklu süreçleri de yansıtır. Örneğin, sentetik yaklaşım zarardan kaçınma davranışını da açıklayabilir. Bu açıklamaya göre zararlı uyaranlar tespit edilmek üzere öncelikli işleme alınmalı ve böylece organizma bu uyaranlardan uzaklaşmalıdır. Ödül/yaklaşma ve zarar/kaçınma süreçlerine yönelik ilkel sinir devreleri, evrimsel tarihin erken dönemlerinde birbirinden ayrılmıştır ve bu da olumlu ve olumsuz değer içerikli uyaranlara ilişkin farklı bulgulara yönelik süreç temelli bir açıklama sunar. Benzer şekilde sentetik yaklaşım, nesnelerin farklı özelliklerine olan duyarlılığın neden eylem bağlamına bağlı olduğunu da açıklar (Bekkering ve Neggers, 2002; Craighero ve ark., 1999; Fagioli ve ark., 2007; Welsh ve Pratt, 2008). Bu yaklaşıma göre bağlam, eylem merkezli parietal dizilerden hangisinin belirli bir anda seçici olarak güçlendirildiğini kendine özgü dış dünya temsiliyle belirler.
Deneysel psikoloji ve sinirbilimin en büyük bilmecelerinden biri, bu bilgi akışlarının başlangıçtaki duyusal alanlardan nasıl dallanıp ayrıldığı ve ardından eyleme dönüşmek üzere nasıl birleştiğidir. Davranışa dayalı gözlemler sayesinde genellikle aynı anda yalnızca bir hedefe yönelik hareket gerçekleştirildiği, ancak birden fazlasının eşzamanlı olarak temsil edilebileceği bilinmektedir (örn. Cisek ve Kalaska, 2005). Evrimsel adaptasyonlar bölümünde tartıştığımız üzere, her bir eylem sistemi için (örneğin el ve göz hareketleriyle desteklenen eylemlerde) tek bir eylem oluşturacak şekilde çözüme ulaşan, “kazanan hepsini alır” tarzı bir dinamikle (“winner-take-all” dynamics) örtüşen paralel ve rekabetçi bir yapıyı savunuyoruz. Elbette, bunun nasıl gerçekleştiğine dair detayların büyük kısmı hala açıklanmamış ve bu makalenin kapsamı dışındadır. Burada vurgulamak istediğimiz, karmaşık duyusal bilginin eyleme nasıl dönüştüğünü anlamamıza yardımcı olan sentetik yaklaşımdır. Çıkaracağımız temel sonuç ise “dikkat” gibi her şeye uydurulmaya çalışılan genel geçer kavramlara başvurmanın veya onları uygulamaya çalışmanın bilimsel ilerlemeyi sekteye uğratmasıdır. Bu nedenle, kişinin “fiziksel olarak belirgin bir uyarana daha fazla dikkat gösterdiğini” söylemek yerine, fiziksel belirginliğin nasıl olup da daha etkili bilgi işleme ve davranışa dönüştüğünü açıklayan mekanizmayı anlamaya çalışmak gerekir. Benzer şekilde, ödüllendirici uyaranların “daha fazla dikkat gerektirdiğini” öne sürmek yerine, belirli bir ödülün belirli bir hedefle nasıl ilişkilendirildiğini ve bilişsel sistemin/beynin bu ilişkiyi saniyenin çok kısa bir diliminde nasıl hatırlayıp sonraki denemelerde davranışı nasıl yönlendirdiğini açıklamak gerekir. Görsel bilgiyi eylem için seçilen bir konumda sunulduğunda neden ve nasıl güçlendiğini deneylerle ortaya koymak, bulguyu “sadece dikkat” olarak nitelendirmekten daha bilgilendiricidir. Bu nedenle, anlaşılabilen mekanizmalara yönelmek ve bilişsel alanlarda ilgilenilen davranışı yeniden oluşturmaya çalışmak gerekir. Bu yaklaşım başarılı olursa, hem açıklanacak olgu (explanandum) hem de açıklayıcı mekanizmayı (explanans) tanımlarken dikkat gibi tanımsız kavramlara gerek kalmayacaktır.
Önerdiğimiz sentetik yaklaşım indirgemeci gibi algılanabilir, ancak bu yaklaşım sinirsel açıklamaları işlevsel açıklamalara üstün kılmamasıyla önem arz eder. Gerçek bir bilişsel sinirbilim, iyi teorilerin hem sinirsel hem de işlevsel kısıtlamaları dikkate alması gerektiği fikrine dayanır; böylece sinirsel ve işlevsel teoriler birbiriyle çelişmez. Ayrıca sinirsel açıklamaların daha temel veya nedensel olduğu yönündeki eğilimleri de mantıksal olarak desteklemez. Örneklerimizin birçoğu sinirsel bulgu ve açıklamalara atıfta bulunsa ve evrimsel muhakememiz ağırlıklı olarak sinirsel terimlerle ifade edilmiş olsa da sinirsel açıklamaların işlevsel olanlara üstünlüğünü savunmuyor ve her iki yaklaşımdan da kanıtlar sunmaya özen gösteriyoruz. Öte yandan, bilişsel psikoloji ve sinirbilim alanlarında iyi teorilerin, gözlemlenen bir olgunun (explanandum) altında yatan mekanizmalar (explanans) aracılığıyla nasıl ortaya çıktığına dair test edilebilir hipotezlerden doğduğunu tümüyle kabul ediyoruz. Böylece bu mekanizmaların sinirsel mi yoksa işlevsel kavramlarla mı tanımlandığı ise ikinci planda kalmaktadır. Ana akımın aksine sentetik yaklaşım, bir olguyu parçalara ayırmak yerine iyi anlaşılmış temel mekanizmalar üzerinden onu yeniden inşa etmeyi gerektirir. Bizim beklentimiz, bu sentetik/yapılandırmacı (constructivist) yaklaşımın nihayetinde, açıklamayı hedeflediğimiz olguları tanımlama biçimlerimizin aslında yanıltıcı olduğunu ortaya çıkaracağı yönündedir. Özellikle de dikkat kavramı için bunun geçerli olduğunu düşünüyoruz. Dolayısıyla, James’in (1890) iddiasının aksine, hiç kimsenin dikkatin tam olarak ne olduğunu bilmediğini ve asla da bilemeyeceğini savunuyoruz.
“Ana akımın aksine sentetik yaklaşım, bir olguyu parçalara ayırmak yerine iyi anlaşılmış temel mekanizmalar üzerinden onu yeniden inşa etmeyi gerektirir.”
Dipnotlar:
[1] William James’in dikkat kavramına yaklaşımının esasen fenomenoloji ile ilgili olduğunu belirtmek gerekir. Bu yaklaşıma, modern işlevsel ve sinirsel yaklaşımları ele aldığımız bu makalede daha fazla yer verilmeyecektir.
Referanslar
Aboitiz, F., & Montiel, J. F. (2015). Olfaction, navigation, and the origin of isocortex. Frontiers in Neuroscience, 9, 402.
Allport, A. (1993). Attention and control: Have we been asking the wrong questions? A critical review of twenty-five years. In D. E. Meyer & S. Kornblum (Eds.), Attention and performance XIV (pp. 183–218). MIT Press.
Allport, D. A. (1987). Selection for action: Some behavioral and neurophysiological considerations of attention and action. In H. Heuer & H. F. Sanders (Eds.), Perspectives on perception and action (pp. 395–419). Lawerence Erlbaum.
Andersen, R. A., Snyder, L. H., Bradley, D. C., & Xing, J. (1997). Multimodal representation of space in the posterior parietal cortex and its use in planning movements. Annual Review of Neuroscience, 20, 303–330.
Anderson, B. (2011). There is no such thing as attention. Frontiers in Psychology, 2, 1–8. https://doi.org/10.3389/fpsyg.2011.00246
Anderson, B. A., & Yantis, S. (2012). Value-driven attentional and oculomotor capture during goal-directed, unconstrained viewing. Attention, Perception & Psychophysics, 74, 1644–1653. https://doi.org/10.3758/s13414-012-0348-2
Anderson, B., Laurent, P., & Yantis, S. (2011). Value-driven attentional capture. Proceedings of the National Academy of Sciences of the United States of America, 108, 10367–10371. https://doi.org/10.1073/pnas.1104047108
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16, 437–443.
Baldauf, D., & Deubel, H. (2008). Properties of attentional selection during the preparation of sequential saccades. Experimental Brain Research, 184, 411–425. https://doi.org/10.1007/s00221-007-1114-x
Baldauf, D., Wolf, M., & Deubel, H. (2006). Deployment of visual attention before sequences of goal-directed hand movements. Vision Research, 46, 4355–4374. https://doi.org/10.1016/j.visres.2006.08.021
Barrett, L. F. (2017). How emotions are made: The secret life of the brain. Houghton Mifflin Harcourt.
Bartolomeo, P. (2007). Visual neglect. Current Opinion in Neurology, 20, 381–386.
Basso, M. A., & May, P. J. (2017). Circuits for action and cognition: A view from the superior colliculus. Annual Review of Vision Science, 3, 197–226.
Baumann, M. A., Fluet, M. C., & Scherberger, H. (2009). Context specific grasp movement representation in the macaque anterior intraparietal area. Journal of Neuroscience, 29, 6436–6448.
Beauchamp, M. S., Petit, L., Ellmore, T. M., Ingeholm, J., & Haxby, J. V. (2001). A parametric fMRI study of overt and covert shifts of visuospatial attention. Neuroimage, 14, 310–321.
Bechtel, W. (2008). Mechanisms in cognitive psychology: What are the operations? Philosophy of Science, 75, 983–994.
Bekkering, H., & Neggers, S. F. W. (2002). Visual search is modulated by action intentions. Psychological Science, 13, 370–374.
Bisley, J. W., & Goldberg, M. E. (2010). Attention, intention, and priority in the parietal lobe. Annual Review of Neuroscience, 33, 1–21.
Bradley, F. H. (1886). Is there any special activity of attention? Mind, A Quarterly Review of Psychology of Philosophy, 43, 305–323.
Broadbent, D. (1958). Perception and communication. Pergamon Press.
Buckner, R. L., & Schacter, D. L. (2004). Neural correlates of memory’s successes and sins. In M. S. Gazzaniga (Ed.), The cognitive neurosciences III (pp. 739–752). MIT Press.
Bundesen, C. (1990). A theory of visual attention. Psychological Review, 97, 523–547.
Buneo, C. A., Jarvis, M. R., Batista, A. P., & Andersen, R. A. (2002). Direct visuomotor transformations for reaching. Nature, 416, 632–636.
Busemeyer, J. R., Gluth, S., Rieskamp, J., & Turner, B. M. (2019). Cognitive and neural bases of multi-attribute, multi-alternative, value-based decisions. Trends in Cognitive Sciences. https://doi.org/10.1016/j.tics.2018.12.003
Butler, A. B. (2000). Chordate evolution and the origin of craniates: An old brain in a new head. The Anatomical Record, 261, 111–125.
Cavanaugh, J., & Wurtz, R. H. (2004). Subcortical modulation of attention counters change blindness. Journal of Neuroscience, 24, 11236–11243.
Chapman, C. S., Gallivan, J. P., & Enns, J. T. (2015a). Separating value from selection frequency in rapid reaching biases to visual targets. Visual Cognition, 23, 249–271.
Chapman, C. S., Gallivan, J. P., Wong, J. D., Wispinski, N. J., & Enns, J. T. (2015b). The snooze of lose: Rapid reaching reveals that losses are processed more slowly than gains. Journal of Experimental Psychology: General, 144, 844.
Chapman, C. S., Gallivan, J. P., Wood, D. K., Milne, J. L., Ansari, D., Culham, J. C., & Goodale, M. A. (2014). Counting on the motor system: Rapid action planning reveals the format-and magnitude dependent extraction of numerical quantity. Journal of Vision, 14, 30.
Chapman, C. S., Gallivan, J. P., Wood, D. K., Milne, J. L., Culham, J. C., & Goodale, M. A. (2010). Reaching for the unknown: Multiple target encoding and real-time decision-making in a rapid reach task. Cognition, 116, 168–176.
Christopoulos, V. N., Kagan, I., & Andersen, R. A. (2018). Lateral intraparietal area (LIP) is largely effector-specific in free-choice decisions. Scientific Reports, 8, 8611.
Cisek, P. (2007). Cortical mechanisms of action selection: The affordance competition hypothesis. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 362, 1585–1599.
Cisek, P. (2019). Resynthesizing behavior through phylogenetic refinement. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-019-01760-1
Cisek, P., & Kalaska, J. F. (2005). Neural correlates of reaching decisions in dorsal premotor cortex: Specification of multiple direction choices and final selection of action. Neuron, 45, 801–814.
Cisek, P., & Thura, D. (2018). Neural circuits for action selection. In D. Corbetta & M. Santello (Eds.), Reach-to-grasp behavior: Brain, behavior, and modelling across the life span. Routledge.
Colby, C. L., & Duhamel, J. R. (1996). Spatial representations for action in parietal cortex. Brain Research: Cognitive Brain Research, 5, 105–115.
Comoli, E., Das Neves Favaro, P., Vautrelle, N., Leriche, M., Overton, P. G., & Redgrave, P. (2012). Segregated anatomical input to subregions of the rodent superior colliculus associated with approach and defense. Frontiers in Neuroanatomy, 6, 9.
Corbetta, M., & Shulman, G. L. (2011). Spatial neglect and attention networks. Annual Review of Neuroscience, 33, 569–599.
Corbetta, M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Ollinger, J. M., Drury, H. A., Linenweber, M. R., Petersen, S. E., Raichle, M. E., Van-Essen, D. C., & Shulman, G. L. (1998). A common network of functional areas for attention and eye movements. Neuron, 21, 761–773.
Craighero, L., Fadiga, L., Rizzolatti, G., & Umiltà, C. A. (1999). Action for perception: A motor-visual attentional effect. Journal of Experimental Psychology: Human Perception and Performance, 25, 1673–1692.
Cui, H., & Andersen, R. A. (2007). Posterior parietal cortex encodes autonomously selected motor plans. Neuron, 56, 552–559.
Culham, J. C., & Kanwisher, N. G. (2001). Neuroimaging of cognitive functions in human parietal cortex. Current Opinion in Neurobiology, 11, 157–163.
Cutrell, E. B., & Marrocco, R. T. (2002). Electrical microstimulation of primate posterior parietal cortex initiates orienting and alerting components of covert attention. Experimental Brain Research, 144, 103–113. https://doi.org/10.1007/s00221-002-1032-x
Deubel, H., & Schneider, W. X. (1996). Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research, 36, 1827–1837.
Deubel, H., & Schneider, W. X. (2003). Delayed saccades, but not delayed manual aiming movements, require visual attention shifts. Annals of the New York Academy of Science, 1004, 289–296.
Deubel, H., Schneider, W. X., & Paprotta, I. (1998). Selective dorsal and ventral processing: Evidence for a common attentional mechanism in reaching and perception. Visual Cognition, 5, 81–107.
Di Lollo, V. (2018). Attention is a sterile concept; iterative reentry is a fertile substitute. Consciousness and Cognition, 64, 45–49. https://doi.org/10.1016/j.concog.2018.02.005
Erwin, D. H., Laflamme, M., Tweedt, S. M., Sperling, E. A., Pisani, D., & Peterson, K. J. (2011). The Cambrian conundrum: Early divergence and later ecological success in the early history of animals. Science, 334, 1091–1097.
Eysenck, M. W., & Keane, M. T. (2000). Cognitive psychology: A student’s handbook (4th Ed.). Psychology Press.
Fagioli, S., Hommel, B., & Schubotz, R. I. (2007). Intentional control of attention: Action planning primes action related stimulus dimensions. Psychological Research, 71, 22–29.
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., & Posner, M. I. (2002). Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience, 14, 340–347.
Fecteau, J. H., & Munoz, D. P. (2006). Salience, relevance, and firing: A priority map for target selection. Trends in Cognitive Science, 10, 382–390. https://doi.org/10.1016/j.tics.2006.06.011
Finlay, B. L., & Uchiyama, R. (2015). Developmental mechanisms channeling cortical evolution. Trends in Neurosciences, 38, 69–76.
Folk, C. L., Remington, R. W., & Johnston J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal Experimental Psychology: Human Perception & Performance, 18, 1030–1044.
Friese, M., Loschelder, D. D., Gieseler, K., Frankenbach, J., & Inzlicht, M. (in press). Personality and Social Psychology Review.
Gallivan, J. P., & Chapman, C. S. (2014). Three-dimensional reach trajectories as a probe of real-time decision-making between multiple competing targets. Frontiers in Neuroscience, 8, 215.
Gallivan, J. P., Barton, K. S., Chapman, C. S., Wolpert, D. M., & Flanagan, J. R. (2015). Action plan co-optimization reveals the parallel encoding of competing reach movements. Nature Communications, 6, 7428.
Gallivan, J. P., Cavina-Pratesi, C., & Culham, J. C. (2009). Is that within reach? fMRI reveals that the human superior parieto-occipital cortex encodes objects reachable by the hand. Journal of Neuroscience, 29, 4381–4391.
Gallivan, J. P., Chapman, C. S., Gale, D. J., Flanagan, J. R., & Culham, J. C. (2019). Selective modulation of early visual cortical activity by movement intention. Cerebral Cortex.
Gandhi, N. J., & Katnani, H. A. (2011). Motor functions of the superior colliculus. Annual Review of Neuroscience, 33, 205–231.
Gersch, T. M., Kowler, E., & Dosher, B. (2004). Dynamic allocation of visual attention during the execution of sequences of saccades. Vision Research, 44, 1469–1483.
Glazebrook, C. M., Welsh, T. N., & Tremblay, L. (2016). The processing of visual and auditory information for reaching movements. Psychological Research, 80, 757–773.
Gottlieb, J. (2012). Attention, learning, and the value of information. Neuron, 76, 281–295.
Graziano, M. S. (2016). Ethological action maps: A paradigm shift for the motor cortex. Trends in Cognitive Sciences, 20, 121–132.
Grillner, S., Robertson, B., & Stephenson-Jones, M. (2013). The evolutionary origin of the vertebrate basal ganglia and its role in action selection. The Journal of Physiology, 591, 5425–5431.
Grossberg, S. (1973). Contour enhancement, short term memory, and constancies in reverberating neural networks. Studies in Applied Mathematics, 52, 213–257.
Herrero, L., Rodriguez, F., Salas, C., & Torres, B. (1998). Tail and eye movements evoked by electrical microstimulation of the optic tectum in goldfish. Experimental Brain Research, 120, 291–305.
Hoffman, J. E., & Subramaniam, B. (1995). The role of visual attention in saccadic eye movements. Perception & Psychophysics, 57, 787–795.
Hommel, B. (2010). Grounding attention in action control: The intentional control of selection. In B. J. Bruya (Ed.), Effortless attention: A new perspective in the cognitive science of attention and action (pp. 121–140). MIT Press.
Hommel, B. (2019a). Pseudo-mechanistic explanations in psychology and cognitive neuroscience. Submitted manuscript.
Hommel, B. (2019b). Affect and control: a conceptual clarification. Submitted manuscript.
Hommel, B., & Colzato, L. S. (2015). Learning from history: The need for a synthetic approach to human cognition. Frontiers in Psychology, 6, 1–4. https://doi.org/10.3389/fpsyg.2015.01435
Hommel, B., & Wiers, R. W. (2017). Towards a unitary approach to human action control. Trends in Cognitive Sciences, 21, 940–949.
Howard, L. A., & Tipper, S. P. (1997). Hand deviations away from visual cues: Indirect evidence for inhibition. Experimental Brain Research, 113, 144–152.
Husain, M., & Kennard, C. (1996). Visual neglect associated with frontal lobe infarction. Journal of Neurology, 243, 652–657.
James, W. (1890). The principles of psychology. Henry Holt.
Johnson, P. B., Ferraina, S., Bianchi, L., & Caminiti, R. (1996). Cortical networks for visual reaching: Physiological and anatomical organization of frontal and parietal arm regions. Cerebral Cortex, 6, 102–119.
Jones, M. R., Grillner, S., & Robertson, B. (2009). Selective projection patterns from subtypes of retinal ganglion cells to tectum and pretectum: distribution and relation to behavior. Journal of Comparative Neurology, 517, 257–275.
Kaas, J. H. (2017). The evolution of mammalian brains from early mammals to present-day primates. In S. Watanabe (Ed.), Evolution of the brain, cognition, and emotion in vertebrates (pp. 59–80). Springer.
Kaas, J. H., & Stepniewska, I. (2016). Evolution of posterior parietal cortex and parietal-frontal networks for specific actions in primates. Journal of Comparative Neurology, 524, 595–608.
Kahneman, D. (1973). Attention and effort. Prentice-Hall.
Kahneman, D., & Tversky, A. (1979). Prospect Theory: An Analysis of Decision under Risk. Econometrica, 47, 263–292.
Kalaska, J. F., & Crammond, D. J. (1995). Deciding not to GO: Neuronal correlates of response selection in a GO/NOGO task in primate premotor and parietal cortex. Cerebral Cortex, 5, 410–428.
Kardamakis, A. A., Saitoh, K., & Grillner, S. (2015). Tectal microcircuit generating visual selection commands on gaze-controlling neurons. Proceedings of the National Academy of Sciences of the United States of America, 112, E1956–1965.
Katz, L. N., Yates, J. L., Pillow, J. W., & Huk, A. C. (2016). Dissociated functional significance of decision-related activity in the primate dorsal stream. Nature, 535, 285–288.
Khan, A. Z., Song, J. H., & McPeek, R. M. (2011). The eye dominates in guiding attention during simultaneous eye and hand movements. Journal of Vision, 11, 9. https://doi.org/10.1167/11.1.9
Klaes, C., Westendorff, S., Chakrabarti, S., & Gail, A. (2011). Choosing goals, not rules: deciding among rule-based action plans. Neuron, 70, 536–548.
Kowler, E., Anderson, E., Dosher, B., & Blaser, E. (1995). The role of attention in the programming of saccades. Vision Research, 35, 1897–1916.
Krauzlis, R. J., Bollimunta, A., Arcizet, F., & Wang, L. (2014). Attention as an effect not a cause. Trends in Cognitive Science, 18, 457–464. https://doi.org/10.1016/j.tics.2014.05.008
Lacalli, T. C. (1996). Frontal eye circuitry, rostral sensory pathways and brain organization in amphioxus larvae: Evidence from 3D reconstructions. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 351, 243–263.
Lacalli, T. C. (2018). Amphioxus, motion detection, and the evolutionary origin of the vertebrate retinotectal map. Evolutionary Development Biology, 9.
Lamb, T. D. (2013). Evolution of phototransduction, vertebrate photoreceptors and retina. Progress in Retinal Eye Research, 36, 52–119.
Markov, N. T., Ercsey-Ravasz, M. M., Ribeiro Gomes, A. R., Lamy, C., Magrou, L., Vezoli, J., . . . Kennedy, H. (2014). A weighted and directed interareal connectivity matrix for macaque cerebral cortex. Cerebral Cortex, 24, 17–36.
Milne, J. L., Chapman, C. S., Gallivan, J. P., Wood, D. K., Culham, J. C., & Goodale, M. A. (2013). Connecting the dots: Object connectedness deceives perception but not movement planning. Psychological Science, 24, 1456–1465.
Moher, J., Anderson, B. A., & Song, J. H. (2015). Dissociable effects of salience on attention and goal-directed action. Current Biology, 25, 2040–2046. https://doi.org/10.1016/j.cub.2015.06.029
Mole, C. (2011). Attention is cognitive unison: An essay in philosophical psychology. Oxford University Press.
Moore, T., & Fallah, M. (2001). Control of eye movements and spatial attention. Proceedings of the National Academy of Sciences of the United States of America, 98, 1273–1276. https://doi.org/10.1073/pnas.021549498021549498
Mountcastle, V. B., Lynch, J. C., Georgopoulos, A. P., Sakata, H., & Acuna, C. (1975). Posterior parietal association cortex of the monkey: command functions for operations within extrapersonal space. Journal of Neurophysiology, 38, 871–908.
Muller, J. R., Philiastides, M. G., & Newsome, W. T. (2005). Microstimulation of the superior colliculus focuses attention without moving the eyes. Proceedings of the National Academy of Sciences of the United States of America, 102, 524–578.
Murray, E. A., Wise, S. P., & Graham, K. S. (2017). The evolution of memory systems: Ancestors, anatomy, and adaptations. Oxford University Press.
Mysore, S. P., & Knudsen, E. I. (2011). The role of a midbrain network in competitive stimulus selection. Current Opinion in Neurobiology, 21, 653–660.
Nakamura, H., Kuroda, T., Wakita, M., Kusunoki, M., Kato, A., Mikami, A., Sakata, H., & Itoh, K. (2001). From three-dimensional space vision to prehensile hand movements: the lateral intraparietal area links the area V3A and the anterior intraparietal area in macaques. Journal of Neuroscience, 21, 8174–8187.
Navon, D. (1984). Resources—a theoretical soup stone? Psychological Review, 91, 216–234.
Neyedli, H., & Welsh, T. N. (2012). The processes of facilitation and inhibition in a cue-target paradigm: Insight from movement trajectory deviations. Acta Psychologica, 139, 159–165.
Neyedli, H. F., & Welsh, T. N. (2015a). Experience and net worth affects optimality in a motor decision task. Motor Control, 19, 75–89.
Neyedli, H. F., & Welsh, T. N. (2015b). The prioritization of probability over value in action selection. Quarterly Journal of Experimental Psychology, 68, 261–283.
Nobre, A. C., Sebestyen, G. N., & Miniussi, C. (2000). The dynamics of shifting visuospatial attention revealed by event-related potentials. Neuropsychologia, 38, 964–974.
Nummela, S. U., & Krauzlis, R. J. (2010). Inactivation of primate superior colliculus biases target choice for smooth pursuit, saccades, and button press responses. Journal of Neurophysiology, 104, 1538–1548.
Paré, M., & Wurtz, R. H. (2001). Progression in neuronal processing for saccadic eye movements from parietal cortex area lip to superior colliculus. Journal of Neurophysiology, 85, 2545–2562.
Pashler, H., & Johnston, J. C. (1998). Attentional limitations in dual-task performance. In H. Pashler (Ed.), Attention (pp. 155–189). Psychology Press.
Pesaran, B., Nelson, M. J., & Andersen, R. A. (2008). Free choice activates a decision circuit between frontal and parietal cortex. Nature, 453, 406–409.
Peterson, M. S., Kramer, A. F., & Irwin, D. E. (2004). Covert shifts of attention precede involuntary eye movements. Perception & Psychophysics, 66, 398–405.
Posner, M. I., & Dehaene, S. (1994). Attentional networks. Trends in Neuroscience, 17, 75–79.
Posner, M. I., Cohen, Y., & Rafal, R. D. (1982). Neural systems control of spatial orienting. Philosophical Transactions of the Royal Society of London, Series B: Biological Sciences, 298, 187–198.
Posner, M. I., Rafal, R. D., Choate, L. S., & Vaughan, J. (1985). Inhibition of return—Neural basis and function. Cognitive Neuropsychology, 2, 211–228.
Pratt, J., & Abrams, R. A. (1994). Action-centred inhibition: effects of distractors on movement planning and execution. Human Movement Science, 13, 245–254.
Ptak, R. (2012). The frontoparietal attention network of the human brain: action, saliency, and a priority map of the environment. Neuroscientist, 18, 502–515.
Rizzolatti, G., & Luppino, G. (2001). The cortical motor system. Neuron, 31, 889–901.
Rizzolatti, G., Riggio, L., & Sheliga, B. M. (1994). Space and selective attention. In C. Umilta & M. Moscovitch (Eds.), Attention and performance XV. MIT Press.
Rizzolatti, G., Riggio, L., Dascola, I., & Umilta, C. (1987). Reorienting attention across the horizontal and vertical meridians: evidence in favor of a premotor theory of attention. Neuropsychologia, 25, 31–40.
Robinson, D. L., Goldberg, M. E., & Stanton, G. B. (1978). Parietal association cortex in the primate: sensory mechanisms and behavioral modulations. Journal of Neurophysiology, 41, 910–932.
Saitoh, K., Menard, A., & Grillner, S. (2007). Tectal control of locomotion, steering, and eye movements in lamprey. Journal of Neurophysiology, 97, 3093–3108.
Sapir, A., Soroker, N., Berger, A., & Henik, A. (1999). Inhibition of return in spatial attention: direct evidence for collicular generation. Nature Neuroscience, 2, 1053–1054.
Scherberger, H., & Andersen, R. A. (2007). Target selection signals for arm reaching in the posterior parietal cortex. Journal of Neuroscience, 27, 2001–2012. https://doi.org/10.1523/JNEUROSCI.4274-06.2007
Sheliga, B. M., Riggio, L., & Rizzolatti, G. (1995). Spatial attention and eye movements. Experimental Brain Research, 105, 261–275.
Snyder, L. H., Batista, A. P., & Andersen, R. A. (1997). Coding of intention in the posterior parietal cortex. Nature, 386, 167–170.
Snyder, L. H., Batista, A. P., & Andersen, R. A. (2000). Intention-related activity in the posterior parietal cortex: a review. Vision Research, 40, 1433–1441.
Snyder, L. H., Grieve, K. L., Brotchie, P., & Andersen, R. A. (1998). Separate body- and world-referenced representations of visual space in parietal cortex. Nature, 394, 887–891.
Song, J. H., & McPeek, R. M. (2010). Roles of narrow- and broad-spiking dorsal premotor area neurons in reach target selection and movement production. Journal of Neurophysiology, 103, 2124–2138. https://doi.org/10.1152/jn.00238.2009
Song, J. H., & McPeek, R. M. (2015). Neural correlates of target selection for reaching movements in superior colliculus. Journal of Neurophysiology, 113, 1414–1422. https://doi.org/10.1152/jn.00417.2014
Song, J. H., & Nakayama, K. (2006). Role of focal attention on latencies and trajectories of visually guided manual pointing. Journal of Vision, 6, 982–995.
Song, J. H., & Nakayama, K. (2008). Target selection in visual search as revealed by movement trajectories. Vision Research, 48, 853–861.
Song, J. H., Rafal, R. D., & McPeek, R. M. (2011). Deficits in reach target selection during inactivation of the midbrain superior colliculus. Proceedings of the National Academy of Sciences of the United States of America, 108, E1433–1440. https://doi.org/10.1073/pnas.1109656108
Stein, J. F. (1992). The representation of egocentric space in the posterior parietal cortex. Behavioral and Brain Sciences, 15, 691–700.
Striedter, G. F. (2005). Principles of brain evolution. Sinauer.
Stuphorn, V., Hoffmann, K. P., & Miller, L. E. (1999). Correlation of primate superior colliculus and reticular formation discharge with proximal limb muscle activity. Journal of Neurophysiology, 81, 1978–1982.
Suchman, R. G., & Trabasso, T. (1966). Color and form preference in young children. Journal of Experimental Child Psychology, 3, 177–187.
Thura, D., & Cisek, P. (2014). Deliberation and commitment in the premotor and primary motor cortex during dynamic decision making. Neuron, 81, 1401–1416.
Tipper, S. P., Lortie, C., & Baylis, G. C. (1992). Selective reaching: Evidence for action-centred attention. Journal of Experimental Psychology: Human Perception and Performance, 18, 891–905.
Tipper, S. P., Meegan, D., & Howard, L. A. (2002). Action-centred negative priming: Evidence for reactive inhibition. Visual Cognition, 9, 591–614.
Wang, X. J. (2002). Probabilistic decision making by slow reverberation in cortical circuits. Neuron, 36, 955–968.
Weir, P. L., Weeks, D. J., Welsh, T. N., Elliott, D., Chua, R., Roy, E. A., & Lyons, J. (2003). Action-centred distractor effects in discrete control selection. Experimental Brain Research, 149, 207–213.
Welsh, T. N. (2011). The relationship between attentional capture and deviations in movement trajectories in selective reaching tasks. Acta Psychologica, 137, 300–308.
Welsh, T. N., & Elliott, D. (2004). Movement trajectories in the presence of a distracting stimulus: Evidence for a response activation model of selective reaching. Quarterly Journal of Experimental Psychology– Section A, 57, 1031–1057.
Welsh, T. N., & Pratt, J. (2008). Actions modulate attentional capture. Quarterly Journal of Experimental Psychology, 61, 968–976.
Welsh, T. N., & Zbinden, M. (2009). Fitts’ Law and action-centred reference frames in selective reaching: The “proximity-to-hand” effect revisited. Motor Control, 13, 100–112.
Welsh, T. N., Neyedli, H. F., & Tremblay, L. (2013). Refining the time course of facilitation and inhibition in attention and action. Neuroscience Letters, 554, 6–10.
Wise, S. P., Boussaoud, D., Johnson, P. B., & Caminiti, R. (1997). Premotor and parietal cortex: corticocortical connectivity and combinatorial computations. Annual Review of Neuroscience, 20, 25–42.
Wispinski, N. J., Gallivan, J. P., & Chapman, C. S. (in press). Models, movements, and minds: bridging the gap between decision making and action. Annals of the New York Academy of Sciences.
Wispinski, N. J., Truong, G., Handy, T. C., & Chapman, C. S. (2017). Reaching reveals that best-versus-rest processing contributes to biased decision making. Acta Psychologica, 176, 32–38.
Wu, W. (2014). Attention. Routledge.
Yantis, S. (1998). Control of visual attention. In H. Pashler (Ed.), Attention (pp. 223–256). Psychology Press.
Yoxon, E., Constable, M. D., & Welsh, T. N. (2019). Probing the time-course of facilitation and inhibition in gaze cueing of attention. Attention, Perception, & Psychophysics.
