Özgün adı: “Is There a ‘Social’ Brain? Implementations and Algorithms”
Öne Çıkanlar
Psikoloji ve sinirbilimde temel bir soru, sosyal davranışın sosyal olarak özelleşmiş (socially-specific) sistemler tarafından ne ölçüde desteklendiği ya da sosyal olmayan (non-social) diğer bilişsel, algısal ve motor becerilerle ne derecede paylaşıldığıdır.
Farklı açıklama seviyelerinde, sosyal açıdan özelleşmiş olabilecek bir süreç öneriyoruz. Bu yaklaşım, ayna nöronların (mirror neurons) (bkz. Sözlük) sosyal bağlamlardaki rolünün ve sosyal öğrenmenin benzersiz bir şekilde “sosyal” olup olmadığının netleştirilmesine yardımcı olabilir. Deneysel tasarım, sosyal özgüllüğün (social specificity) farklı düzeylerde mümkün olduğuna dair bir anlayışla yönlendirilmelidir.
Sosyal davranışı türler arasında incelemek, farklı uygulamalar ve algoritmalar hakkında insanlara benzersiz ipuçları verebilir. Örneğin kanıtlar aynı noktada birleşerek, beyinde anterior singulat girusun sosyal uygulamaların işlenmesinde çok önemli olduğunu ve “zihin teorisinin” (theory of mind) (bkz. Sözlük) varsayılan bir sosyal algoritma olduğunu vurgulamaktadır.
Özet
Psikoloji ve sinirbilimde temel bir soru, sosyal davranışın sosyal olarak özelleşmiş sistemler tarafından ne ölçüde desteklendiği ya da sosyal olmayan diğer bilişsel, algısal ve motor becerilerle ne derecede paylaşıldığıdır.
Bu çalışmada, bilgi işlemenin farklı düzeylerde “sosyal” veya “sosyal olmayan” olarak anlaşılabileceğini öne sürmek için, Marr’ın (1982) çerçevesini insanlar, maymunlar ve kemirgenler üzerindeki araştırmalara uyguluyoruz. Süreçlerin sosyal olarak uygulama düzeyinde ve / veya algoritmik düzeyde özelleşebileceğini ve sosyal davranışın hedefini değiştirmenin sosyal özgüllüğü de değiştirebileceğini savunuyoruz. Bu çerçeve, türler arasındaki sosyal davranışın doğasına ilişkin önemli yeni bilgiler sağlayabilir, daha fazla entegrasyonu kolaylaştırabilir, yeni teorik ve ampirik yaklaşımlara ilham verebilir.
Anahtar kelimeler: sosyal biliş, sosyal davranış, bilişsel süreç, beyin devresi, hücresel, işlemlemesel, insanlar, primatlar, kemirgenler
Marr’ın Gözüyle Sosyal Özgüllük
Çoğu davranış sosyal bağlamda ortaya çıkar. Sosyal davranışlar, tek hücreli mikroorganizmalardan [1] kemirgenlere [2], balıklara [3] ve primatlara [4] kadar şaşırtıcı derecede geniş bir tür dizisi boyunca sergilenir. Bununla birlikte, psikoloji ve sinirbilim için temel bir soru, bilgiyi sosyal olarak özelleşmiş bir şekilde işleyen bilişsel süreçlerin, beyin bölgelerinin, devrelerin veya hücrelerin olup olmadığıdır. Yani, “sosyal olmayan” bilişsel, motor ve algısal yetenekler için gerekli olandan farklı şekilde, yalnızca sosyal durumlarda çevrimiçi hale gelen süreçler mi söz konusudur?
Bilişsel ve sinirsel süreçlerin sosyal olarak özelleşip özelleşmediğini anlamak, test etmek ve bu bağlamda bir çerçeve sağlamak için Marr’ın seviyelerinin (bkz. Sözlük) öncü fikrinden yararlanıyoruz [5]. Algoritmik düzeyde sosyal olarak kabul edilebilecek — sosyal olmayan bir alanda işlenenden farklı olan belirli bir algoritma veya kurallı kodlar — ve / veya uygulama düzeyinde aynı algoritmanın kullanıldığı, ancak farklı bir beyin alanı, devresi veya hücrede işlendiği bir süreç olduğunu savunuyoruz. Ayrıca işbirliği / rekabet sırasında olduğu gibi bilgi işleme sisteminin (işlemleme düzeyi — computation level) sosyal hedefini değiştirmenin, diğer düzeylerdeki sosyal özelleşmeyi etkileyebileceğini ileri sürüyoruz. Sosyal bağlamlarda bilgi işlemeyi incelerken, bu tanımlama seviyeleri genellikle gözden kaçar. Bunun bilişsel veya sinirsel süreçlerin özelleşip özelleşmediği konusunda yanlış sonuçlara yol açabileceğini iddia ediyor ve basit çağrışımının ötesinde “sosyal beyin” ifadesine daha incelikli bir yaklaşım çağrısında bulunuyoruz.
Marr’ın Çerçevesi: İşlemleme, Algoritma ve Uygulama
Marr’ın çerçevesi [5], bir bilgi işleme sistemini anlamak için, birden çok açıklama düzeyini dikkate almanın — işlemlemesel (computational), algoritmik ve uygulamalı (Şekil 1) [6] — çok önemli olduğunu savunur. En yüksek açıklama seviyesi olan işlemleme, bir sistemin nedenini veya gerçekleştirmek istediği hedefi açıklar. Örneğin kuşların uçuşunu anlamak istiyorsak, bunu “sadece tüyleri inceleyerek” yapamayız [5]. Öncelikle kuşun amacının uçmak olduğunu bilmeliyiz. İkinci seviye ise algoritmiktir — beyin belirli bir operasyon için hangi kuralları uygular? İşte bu seviyede, kuşun kanatlarını çırpması söz konusudur. Son olarak, uygulamaya yönelik seviye, yani beynin belirli bir operasyonu ‘nasıl’ başardığıdır. Bir kuş için bu, onun tüylerinin olmasıdır.
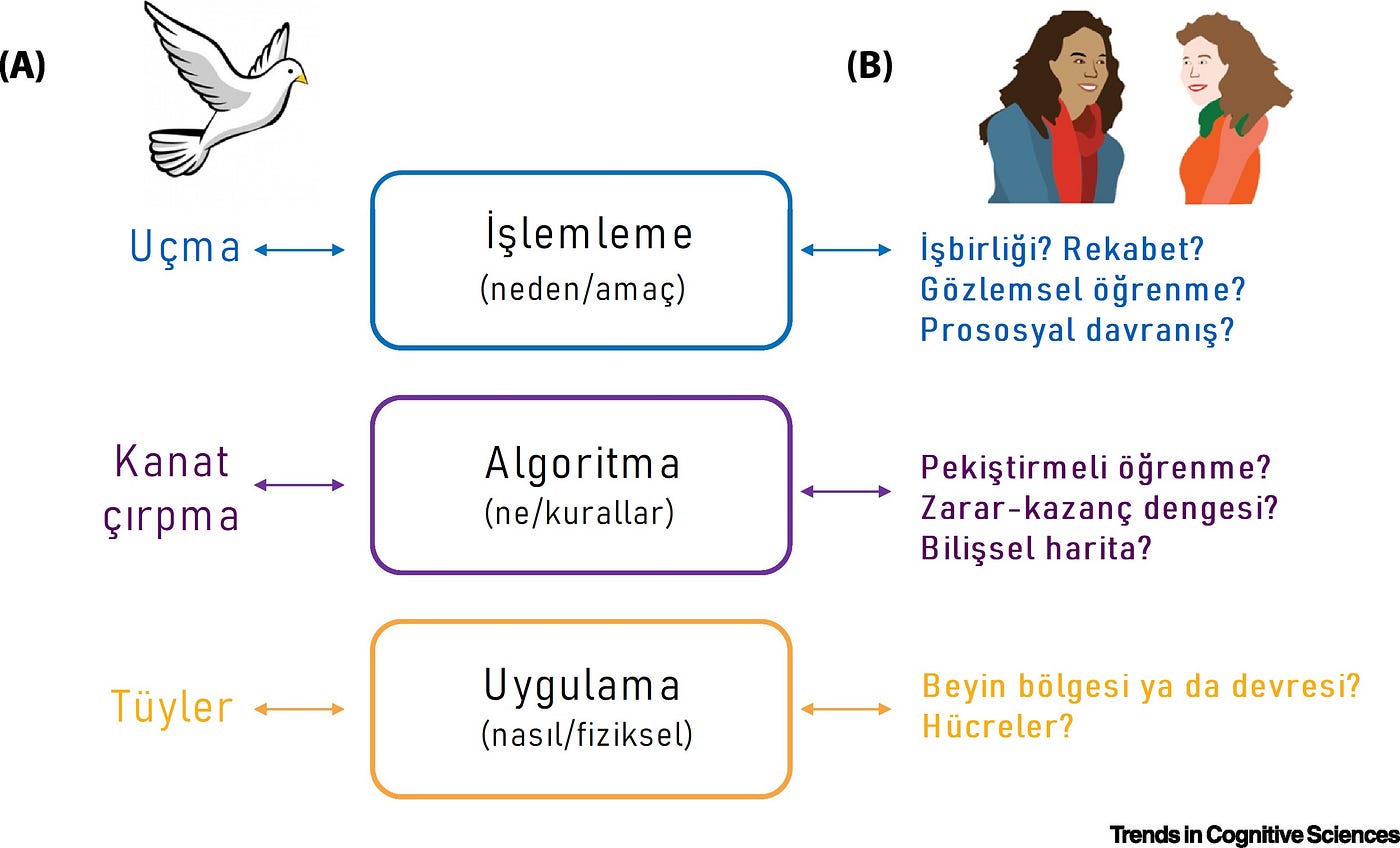 Şekil 1: Marr’ın Sosyal ve Sosyal Olmayan Davranış için Üç Seviyeli Analizi.
Şekil 1: Marr’ın Sosyal ve Sosyal Olmayan Davranış için Üç Seviyeli Analizi.
(A) Bir kuşun uçma amacını anlamak istiyorsak, sadece tüylerini inceleyemeyiz. Kuşun uçma amacının (işlemleme), kanatlarını çırparak (algoritma) yaptığı, uçmanın aerodinamiğinin tüylere bağlı olduğunu (uygulama) bilmemiz gerekir. (B) Kuş uçuşu gibi diğer “sosyal olmayan” süreçlerle karşılaştırıldığında belirli sosyal davranışın ne olduğunu anlamak için, sosyal hedefi anlamamız gerektiğini savunuyoruz (İşbirliği yapıyor muyuz, diğer kişiden öğreniyor muyuz veya diğer kişiye ya da gruba yardım ediyor muyuz?). Sonrasında, bunu başardığımız algoritmayı anlamamız gerekiyor. Pekiştirmeli öğrenme (reinforcement learning) (bkz. Sözlük), zarar-kazanç değiş tokuşu ve bilişsel haritalar gibi işlemleme modellerinin nispeten yeni kullanımı, kullanılabilecek bazı algoritma örnekleridir. Son olarak, sosyal sürecin nasıl uygulandığını ve hangi beyin alanlarında, devrelerinde veya hücrede gerçekleştirildiğini bilmemiz gerekir. En önemlisi, sosyal özgüllük olduğu sonucuna varmak için sosyal ve sosyal olmayan süreç arasında, algoritma düzeyinde veya uygulama düzeyinde bir ayrışmanın gerekli olduğunu savunuyoruz. Sosyal özgüllüğü test etmek için deneyler tasarlarken, algoritma veya uygulamadaki ayrışmaları aramalıyız.
Bu teori, sosyal davranışa nasıl uygulanabilir? Sosyal etkileşimlerin işlemleme amacı, temsilcinin benzerlerle işbirliği yapma, onlara bağlı olma veya onlarla rekabet etme gibi doğası ve niyetleri tarafından belirlenir. Algoritmik seviye, yalnızca sosyal etkileşim esnasında uygulanan sosyal veya bilişsel bir sürecin belirli bir modeli olacaktır. Son olarak, uygulama düzeyi, sosyal sürecin gerçekleştirildiği belirli beyin bölgesi, devre veya hücre olacaktır. Düzeylerin sayısı ve bağımsızlıkları tartışılsa da [6,7], Marr’ın teorisi sosyal özgüllüğün farklı düzeylerde tasvir edilebileceğini öne süren önemli bir organizasyon çerçevesi sağlar. En can alıcı nokta ise, bir sürecin sosyal olarak özelleşmiş olarak kabul edilebilmesi için, algoritmik veya uygulamalı düzeylerde sosyal ve sosyal olmayan işlemler arasında bir ayrışma olmasıdır. Ayrıca alternatif, benzer, alan-geneline (domain-general) yönelik süreçler göz ardı edilmelidir.
Marr’ın çerçevesi, sosyal sinirbilimdeki çeşitli tartışmalara yeni bakış açıları sunar. Biz bu çalışmada iki önemli örneği ele alıyoruz. Ayna nöronlar veya ‘ortak değer’ açıklamaları, sosyal bilginin uygulamadaki örtüşme durumuna dayalı olarak (aynı nöron benzer şekilde ateşlenir veya fMRI kan oksijen seviyesine bağlı- BOLD: Blood oxygenation level dependent– aynı yanıt gözlenir) birinci ve üçüncü şahıslarla kodlandığını gösterir. Örnekler arasında kendine ve başkasına dair acı çekme [8,9], para ve sosyal ödül (bkz. Sözlük) [10,11] ve birinin kendi veya diğeri için eylemsel hedefleri gözetmesi [12,13] yer alır. Bu örtüşme, farklı süreçlerin “ortak kodlaması” olarak yorumlanır, yani hem birinci hem de üçüncü şahıs anlayışının veya empatinin meydana geldiği [6] söylenir. Bununla birlikte, aynalama ile ilgili bu tür sonuçlar, sadece uygulama düzeyine atıfta bulunularak çıkarılır. Peki ya kişi bir başkasının durumunu anlamak için farklı bir algoritma kullanılıyorsa? Algoritmik seviyeyi manipüle veya kontrol etmeden, aynalama (mirroring) sosyal özgüllüğün ya yalnızca uygulamada yokluğunu ya da yalnızca belirli bir devre ya da hücrede var olduğunu yansıtabilir. Gerçekten de, başka birinin eylemlerini izlerken sistemin hedef seviyesinin farklı olduğu açıktır. Eylemleri yeniden zorunlu olarak kendimiz üretmeyi hedeflemiyoruz ve bu nedenle uygulamada örtüşme olsa bile, işlevsel ayrışma olması muhtemel. Bununla birlikte, diğer seviyelerin, sosyal biliş ve davranış — eğer varsa — bazında örtüşmenin ne anlama geldiğini anlamak ve bu bağlamda onları manipüle etmek için dikkatlice kontrol edilen tasarımlar kullanılarak ölçülmesi gerekir (Ek 1).
Ek 1
Sosyal Olmayan Kontrol Koşullarının Önemi
Bilişsel veya sinirsel bir sürecin sosyal davranışta seçici bir role sahip olup olmadığını test etmek için bir deney tasarlarken, uygun kontrol koşulları hakkında düşünmek önemlidir. Bu fikir bilim felsefesindeki yanlışlamanın gerekliliğine dair temel ilkelerden kaynaklanmaktadır [91]. Örneğin, belirli bir beyin bölgesinin hem gülen yüzlere hem de parasal ödüle tepki verdiğini bulursak, bu alanın uyaranın ne kadar ödüllendirici olduğu hakkında bilgi kodladığı sonucuna varabilir miyiz? Bu durumda, ilgili beyin bölgesinin başka bir sürece dahil olmadığı gösterilmedi, sadece ortak bir özelliği paylaşan iki süreçte yer aldığı gösterildi. Kemirgenler ve maymunlarda yapılan birçok çalışma, açık “sosyal olmayan” kontrol koşullarını içermektedir [21,42,45,47,50,51]. Maymunlarda kendine ve diğerine ödül işleme çalışmalarında, ödülün ne kendisine ne de akrabasına verilmediğini gördüğü ödül koşulu eklenmiştir [45,63]. Gözlemsel korku koşullandırmasının olduğu kemirgen çalışmalarında, caydırıcı olaylara “genel alan” yanıtını dahil etmemek için, herhangi bir sosyal bağlam olmaksızın daha tipik bir klasik koşullandırma koşulu kullanılmıştır. Zihin teorisi işleme ile ilgili bazı çalışmalarda, zihin teorisi işlemenin özgüllüğünü göstermeye çalışmak için bir “bilgisayar” koşulu veya bir fiziksel nesne koşulu getirilmiştir [65,78, 79].
Ancak bu kontrol koşulları her zaman deneysel tasarımın bir parçası değildir. Bazen, bir sosyal uyaranın sosyalliği dışındaki tüm özelliklerini veya çoğunu paylaşan, yani başka bir kişi veya grupla ilgili veya onlar için olan bir uyarıcı olan, eşit derecede eşleştirilmiş sosyal olmayan bir kontrol oluşturmak çok zor olabilir. Bilgisayar örneğinde, insanlar bilgisayarı insanlaştırıyor olabilir ve bu nedenle uyarıcıyı sosyal bağlamla ilişkilendirmeye devam edebilir, bu nedenle katılımcıların durumu nasıl algıladıklarını kontrol etmek değerli olabilir. Sosyal etkileşimi ima edecek şekilde hareket eden basit geometrik şekillerin sosyal olarak yorumlanabileceği oldukça iyi bilinmektedir [92]. Bu nedenle, sosyal olmayan veya sosyal olmayan bir durum yaratmada merkezi bir faktör, gözlemlenen davranıştan ziyade, kişinin uyaranın sosyal veya sosyal olmadığı hakkında sahip olduğu inançlar gibi görünmektedir. Stanley ve meslektaşlarının [93], inançların ve davranışların uyaranları sosyal olarak algılamadaki rolünü araştırmak için 2 × 2 tasarım kullanan bir çalışmada açıkça gösterilmiştir. Katılımcılar nokta hareketli animasyonları gözlemlemişlerdir. Bu noktaların ya önceden kaydedilmiş insan hareketinden ya da bilgisayarda oluşturuldukları söylenmiştir. Ayrıca, Stanley ve meslektaşları nokta gösteriminin biyolojik olarak makul veya mantıksız olacak biçimde hız profillerini takip edip etmediğini de manipüle etmişlerdir. Katılımcılar, hız profillerinin biyolojik olarak mantıklı olup olmadığına bakılmaksızın, insan hareketini yansıttığı söylendiğinde, ekran parazitiyle karşılaşmıştır. Dolayısıyla bu çalışma, sosyal ve sosyal olmayan deneysel koşulların yaratılmasında uyaranın sosyal ve sosyal olmadığı inancını teşvik etmenin çok önemli olduğu fikrini desteklemektedir.
Bir başka önemli tartışma, sosyal öğrenmenin eşsiz sosyal süreçler gerektirip gerektirmediği veya alan genelindeki ilişkisel öğrenmeden mi kaynaklandığıdır [14, 15, 16]. Aynı çağrışımsal algoritmaların aslında hem kişisel hem de sosyal öğrenme için kullanılabileceğine dair artan kanıtlar vardır [4,17, 18, 19, 20], bunun sosyal özgüllüğün yokluğunu yansıttığı iddia edilebilir. Peki ya uygulama düzeyi ne olacak? Algoritmik düzeyin aksine, hücrelerdeki veya devrelerdeki bazı sosyal öğrenme süreçleri için, kişinin kendi eylemlerinin sonuçlarından öğrenmeye dahil olmayan uygulama düzeyinde ayrışma olduğuna dair artan bir fikir birliği vardır [17,19, 20, 21, 22, 23, 24, 25]. Bu nedenle sosyal öğrenme, algoritma aynı olsa bile, sinirsel uygulama düzeyinde “sosyal olarak özelleşmiş” olabilir.
Bunlar yalnızca birtakım örnekler, fakat amaç sosyal sinirbilim ve psikolojideki tartışmaların Marr’ın öne sürdüğü düzeylerden hangisini irdelediğini dikkate alarak, nasıl ele alınabileceğini vurgulamaktır. Böyle bir fikir, sosyal biliş ve onun sinirsel temeli çalışmalarındaki deneysel tasarımı aydınlatabilir. Bu tür tartışmaları deneysel olarak ele almanın anahtarı, diğer seviyedeki özgüllüğü test etmek için, düzeylerden birinin algoritma veya uygulama olarak sabit tutulduğu deneysel tasarımları kullanmak olacaktır. Örneğin, lezyon ve beyni uyarma yaklaşımları, uygulama düzeyini bozmanın algoritmik düzeydeki etkisini açıkça inceleyebilir. Belirli bir uygulamayı bozabilir ve sosyal veya sosyal olmayan bir algoritmanın değiştirilip değiştirilmediğini test edebiliriz. Algoritmik düzeyde, pekiştirmeli öğrenme süreci gibi ortak bir algoritmayı inceleyebilir ve ardından sosyal olmayan koşullara karşı sosyal bir durumda farklı şekilde uygulanıp uygulanmadığını test edebiliriz. Bu nedenle, farklı analiz düzeylerinin nasıl ayrıştırılacağını düşünmek, özgüllüğü ele almayı amaçlayan deneysel tasarımlar oluşturmada önemlidir. Dahası, amaç sosyal özgüllüğü tanımlamaksa, ek “sosyal olmayan” kontrol koşullarının test edilmesinin kritik olduğunu öne sürüyoruz (Ek 1). Aşağıdaki bölümlerde, bu hipotezi, insanlarda, insan olmayan primatlarda ve kemirgenlerde yapılan araştırmalardan anahtar örnekler kullanarak inceliyoruz.
Seviyeler ve Türler Arasında Sosyal ve Sosyal Olmayan İşleme
Sorulması gereken ilk soru — neden herhangi bir açıklama düzeyinde sosyal özgüllük bulmayı bekleyebiliriz? Evrimsel olarak, hayvan türleri fiziksel ortamlara uyum sağlarlar ve sıklıkla eşleri ile etkileşime giren türler oldukları için aynı zamanda sosyal ortamlara da uyum sağlarlar [26]. Sosyal beyin hipotezi (the social brain hypothesis), sosyal çevrelerde gezinmek için gerekli olan bilişsel becerilerin diğer hayvanlara göre büyük primat beyinlerini şekillendirdiğini [27, 28, 29, 30] ve önceki sosyal zeka hipotezi (the preceding social intelligence hypothesis) ise, sosyal grup yapılarının hayvanlarda daha yüksek zekanın ortaya çıkmasına neden olan evrimsel bir baskı olduğunu savunur [31, 32]. Kemirgenlerde, sosyal bağlamlarda koku alma ve seslendirme, evrimsel uygunluk ile güçlü bir şekilde bağlantılıdır [33, 34]. Bununla birlikte, farklı türlerin beyinlerinin sosyal çevrelerine uyarlanmasının makul olduğunu düşünsek bile, bu sinir sistemlerinin ve süreçlerinin özelleşmesi gerektiği anlamına gelmez.
Dahası, insanların sosyal yaratıklar olduğunu söylemeye gerek kalmasa da, insan olmayan primatlarda ve özellikle kemirgenlerde sosyal davranışın karmaşıklığı ve sınırları geniş çapta tartışılmaktadır [2, 4, 35, 36]. Örneğin birçok kişi, insanların başkalarıyla empati kurma ve zihin teorisi gibi sosyal işlemlerle meşgul olduğu konusunda hemfikirken, bu tür süreçler insan olmayan primatlarda ve kemirgenlerde çok daha tartışmalıdır [4, 35, 36, 37]. Bu durum, sosyal özgüllüğü incelerken önemlidir — aynı sosyal biliş ve davranışlar türler arasında paylaşılmazsa, o zaman algoritmaların ve uygulamaların aynı olmasından (korunduğundan) çok, farklı olmasını bekleyebiliriz. Sosyal davranışın karmaşıklığındaki açık farklılıklara ek olarak, beyin alanlarının benzeşikliğinde [38] ve metodolojik yaklaşımlarda da bazı [2, 36] (Ek 2) farklılıklar vardır. Bu metodolojik yaklaşımlar, kemirgenlerdeki tek hücrelerde optogenetikten (bkz. Sözlük) insanlarda tam beyin nörogörüntülemeye kadar deneysel methodlara göre de büyük farklılıklar gösterebilir ve bu nedenle özgüllük tanımı, çözünürlüğün bir işlevi olarak değişebilir. Ancak bu farklılıklara rağmen türler arasında benzer deneysel teknikler kullanıldığından, belki de en kolay şekilde, ödüllendirici ya da cezalandırıcı uyarıcıların işlenmesine dair paralellikler kurulabilir [4, 39, 40]. Bu nedenle, şu sorular sorulabilir : Sosyal davranışları destekleyen belirli sinir devreleri ve hücreleri var mı? Farklı açıklama seviyelerinde görünüyorlar mı? Uygulama ve algoritmik düzeylerde sosyal özgüllükte net ayrışmalara dair kanıt var mı?
Ek 2
Türler Arası Karşılaştırmaların Fırsatları ve Zorlukları
Farklı türlerden gelen bulguları yürütmeye ve yorumlamaya çalışırken birçok zorluk vardır. Beyin bölgeleri ve devreleri üzerindeki farklılıklar, deneysel tasarımdaki farklılıklar ve yöntemlerin çözümlenmesindeki farklılıklar en önemlilerindendir. Bu zorluklara rağmen, nihayetinde insan davranışını ve patolojilerini anlamak istiyorsak, karşılaştırmalar yapılabilir ve bu anahtar bir rol oynar. Sosyal davranışları araştırmak için insanlarla çalışmalar yürütürken, onların düşüncelerini ve duygularını sorgulama ve anlamaya çalıştığımız sistemi inceleme avantajına sahibiz. Bununla birlikte, etik olarak fokal lezyonlara neden olamayız ve şu anda tek hücreleri manipüle etmek için çok kesin optogenetik yöntemler kullanamıyoruz. Tersine, kemirgenlerde çok kesin optogenetik yöntemler kullanabiliriz, ancak daha sonra kemirgen davranışının insan sosyal davranışına ne kadar benzer olduğunu sorgulayabiliriz ve kesinlikle onların düşüncelerini ve hislerini sorgulayamayız. Bu durum, sosyal davranışı incelemek için birden fazla yöntemi farklı türler arasında kullanmanın ve yakınsak bilgi elde etmek için bunları en anlamlı şekilde karşılaştırmanın fırsatlarının ve zorluklarının farkında olmanın önemini vurgular. Ayrıca, farklı türlere odaklanan yeni deneysel yaklaşımlara ve araştırma alanlarına ilham vermek için de yararlı olabilir. Örneğin, maymunlarda sosyal davranışta ACCg üzerine yapılan çalışmalar, insanlarda ACCg’nin insan ve kemirgen sosyal davranışındaki önemini doğrulayan daha sonraki birkaç çalışmaya ilham vermiştir [21,42]. Algoritma ve uygulama düzeyinde sosyal özgüllük öneren Marr’ın organizasyon çerçevesi, farklı türlerde kolaylıkla uygulanabilir ve gelecekteki araştırmalarda birden çok düzeydeki bulgular arasında daha büyük bağlantılar kurmamızı sağlayabilir.
Sosyal Olarak “Özelleşmiş bir Uygulama” Olabilir mi?
Uygulama düzeyinde sosyal davranış için belirli sinir sistemlerinin olup olmadığı üzerine ilk sorudan başlarsak, anterior (ön) singulat korteks (ACC) anahtar bir adaydır (Şekil 2). ACC ayrıca, türler arası karşılaştırmalar için idealdir çünkü maymunlarda ve kemirgenlerde deneysel bağlamda nispeten çokça çalışılmıştır [21, 38, 41, 42, 43, 44]. En ilginç şekilde, kanıtlar, ACC içindeki alt bölgeler, özellikle sulkus (ACCs) ve girus (ACCg) arasındaki sosyal ve sosyal olmayan uygulamadaki önemli bölünmelere işaret eder [4, 21, 39, 42, 45, 46] (Şekil 2). Chang ve arkadaşları tarafından yapılan ufuk açıcı bir çalışma [45] ACCg ve ACCs’de sırasıyla sosyal ve kendine yönelik ödül işleme için farklı özelleşme seviyeleri olduğunu göstermiştir. Araştırmacılar, ACCg, ACCs ve orbitofrontal korteksten (OFC) tek birimli aktivite (single-unit activity) ile, deneydeki maymunların kendilerine ödül verme, başka bir maymuna ödül verme veya hiç kimseye ödül vermeme kararı aldığı anda kayıt almıştır. OFC nöronları, ağırlıklı olarak kendilerine ödül verdikleri karar sonuçlarına yanıt vermiş, ACCs nöronları ise kendilerine ödül vermekten vazgeçtikleri kararın sonuçlarını izlemiştir. Aksi bir şekilde, ACCg’deki nöronlar ağırlıklı olarak belirli bir maymunun ödül sonucunu kodlamıştır. Bazı nöronlar, başkalarının ödüllerine özel olarak yanıt verirken (diğeri için atıfta bulundu), başka bir küme, “aynaya benzer”, kendinin ve diğerinin ödüllerini kodlayan (her ikisi için de atıfta bulundu) bir yanıt göstermiştir. Bu sonuçlar, tek tek hücreler düzeyinde bazı sosyal özgüllüklerin olduğunu göstermektedir. Çünkü ACCg’de test edilen nöronlar, başkalarının ödüllerine diğer alanlara göre daha belirgin yanıt vermiştir.
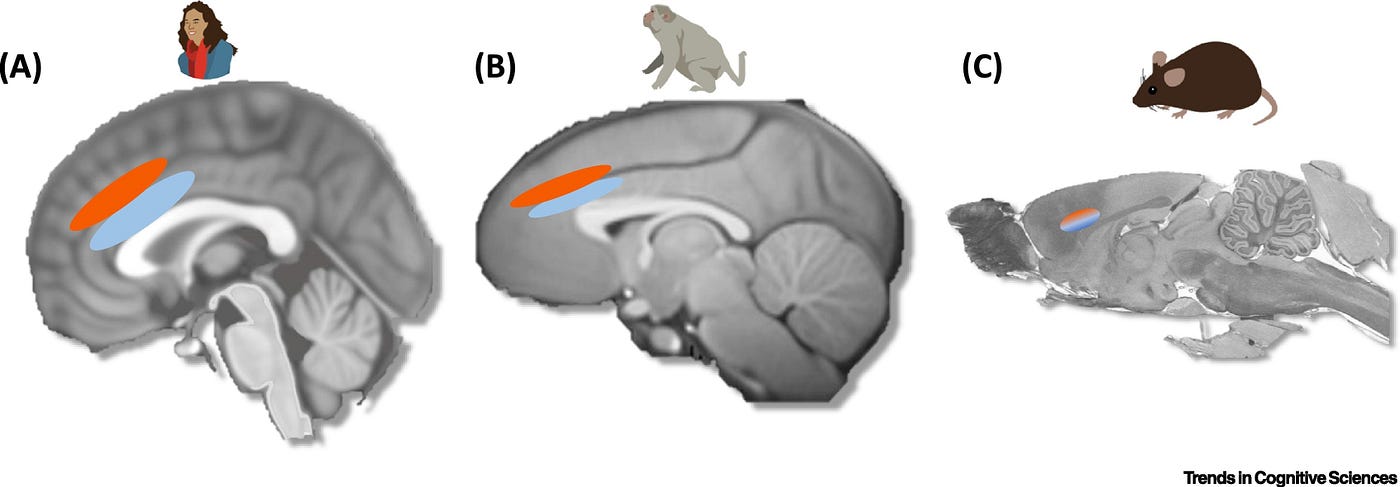 Şekil 2: Anterior Singulat Korteks (ACC) ve Sosyal Olarak Özelleşmiş Uygulama.
Şekil 2: Anterior Singulat Korteks (ACC) ve Sosyal Olarak Özelleşmiş Uygulama.
(A) İnsanlarda, birçok çalışma, ACC’nin giral kısmının (ACCg, açık mavi) tahmin hatalarına ve diğer insanlara verilen ödüllere ya da buna yönelik tahminlere yanıt verdiğini, buna karşılık komşu sulkusun (ACCs, kırmızı) kendisiyle ilgili ödül sinyallerine ve tahmin hatalarına yanıt verdiğini bulmuştur [4, 17, 21, 42, 43]. Daha da önemlisi, ACCs’in tahmin hatalarına yanıt verdiğine ve ödülleri es geçtiğine dair kanıtlar da vardır, bu da söz konusu bölgenin sosyal olarak özelleşmiş bir sinyali işlemek yerine, kendine teslim edilmeyen ödüllere genel bir yanıt işleyebileceğini ima eder [4, 17, 21, 42, 43]. (B) Odak lezyonu ve tek birimli kayıt çalışmalarından elde edilerek birleştirilen kanıtlar, ACCg’nin diğer maymunların ödüllerine dikkat dağıtımı ve davranış seçimi açısından yanıt verdiğini, maymunların ne kendini ne de başkasını ödüllendirmemesinden ziyade başkalarını ödüllendirme tercihi gösterdiklerine işaret etmektedir. Bunun yerine, ACCs’deki nöronların büyük bir kısmı hem kendi ödüllendirdiğinde hem de “hiç kimseyi” ödüllendirmediğinde sinyal vermektedir, bu da söz konusu durumla tutarlıdır [45, 47, 48]. © Kemirgenlerde, sulkus ve girusun net bölünmeleri, insanlarda ve maymunlarda olduğu kadar kolay görülmez, ancak kabaca Cg1 ve Cg2 olarak bilinen alanlara karşılık gelir. Kanıtlar, kemirgenlerdeki ACC’nin hem sıçanın kendisine verilen ayak şoklarına hem de başka bir sıçana verilen şokların gözlemine yanıt veren nöronlar içerdiğini göstermektedir. Önemli olarak, bu aynı nöronlar korku şartlanmasına yanıt vermez [50, 51]. Sıçanda ACC’deki yanıtın kemirgenler ve diğer türler arasındaki türsel farklılıkları mı yoksa beyin evrimine işaret eden farklılıkları mı yansıttığı açık bir sorudur.
Bazı hücreler maymunların ACCg’deki kendi ödülüne yanıt vermesine rağmen, lezyon çalışmaları bir bölgenin işlevinin sosyal davranışın altında olup olmadığına dair nedensel kanıt sağlayabilir. Çarpıcı bir şekilde, ACCg bölgesinin tamamı bozulduğunda, sosyal bilgiye olan dikkat bozulurken, komşu sulkustaki lezyonlar dikkati olduğu gibi sosyal bilgilere bırakır [47]. Bu etkilerin sosyal özgüllüğünü daha da güçlendiren ACCg lezyonları, duygusal uyaranların (bir yılan) veya kontrol nesnelerinin işlenmesini değiştirmemiştir. Benzer şekilde, tüm ACC’yi lezyona uğratan yakın tarihli bir çalışma, uyaranların kendini ödüllendirmediği ancak diğerlerini ödüllendirdiği öğrenmede belirgin bir bozulma gözlemlemiştir [48]. Bunun aksine, önceden öğrenilen uyaranlarla değil, kendini ve diğerini ödüllendirme tercihleri korunmuştur. Bu etkilerin büyük ölçüde ACCg lezyonundan kaynaklanıp kaynaklanmadığı test edilmelidir [49]. Öte yandan, uygulama düzeyinde, ACC’ye verilen zararın seçici olarak sosyal öğrenmeyi etkilediğine dair net kanıtlar sunmuşlardır (Şekil 2).
Kemirgenlerde yapılan araştırmalar, ACC’nin sosyal davranışta anahtar rolüne de işaret etmiştir [44, 50, 51, 52, 53, 54]. Cg1 ve Cg2’nin insan ACC’nin bir homologu olabileceği düşünülse de, kemirgenlerde sulkal ve giral kısımlar arasında bir bölünme insanlarda ve primatlarda olduğu kadar açık değildir [38]. Birbirine yaklaşan kanıtlar, kemirgen ACC’nin sosyal bağlamlarda ödül ve acı işlemeyle bağlantılı olduğuna işaret ediyor. Örneğin, sıçanlarda yapılan çalışmalar, başkalarına zarar veren eylemlerden kaçındığını ve bu etkinin ACC inaktivasyonu ile ortadan kaldırıldığını ileri sürmüştür [51]. Benzer şekilde, ACC inaktivasyonu, klasik koşullanmayı olduğu gibi bırakırken gözlemsel korku öğrenimini bozmaktadır [55]. Aksi bir biçimde, amigdala lezyonları ise hem gözlemsel öğrenmeyi hem de klasik korku öğrenmeyi bozarak kendi başına öğrenmede bir rol olduğunu düşündürür. Makaklardaki [45] çalışmayla uyumlu olarak, kemirgen ACC’deki nöronların daha büyük bir oranı, sadece kendisine olana (%12) göre spesifik olarak diğerlerinin ağrısına (%27) ve hem başkalarının hem de kendi ağrısına (%34) daha çok yanıt vermiştir [50]. Dahası, ACC bölgesinin devre dışı bırakılması, sosyal acı (bkz. Sözlük) bağlamında donma tepkilerini bozmakta, ancak sosyal olmayan korkularda bu etkiyi göstermemektedir. Bu çalışmaya [50] korku durumunun dahil edilmesi ve karşılaştırılması, makaklarla yapılan çalışmada ise “hiç kimseye” ödül vermeme koşulu [45], “her ikisi” durumu için önemli bir kontrol sağlar (Şekil 3); uygulama düzeyinde kendisiyle ve diğeriyle örtüşme söz konusudur. Bir nöronun veya beyin bölgesinin özellikle “acı” ya da “ödül” ile ilgili olup olmadığını belirlemek, bunun genel olarak itici ve iştah açıcı uyarıcıları işlemeye dahil olduğunu dışarıda bırakmayı gerektirir (Ek 1).
 Şekil 3: Uygulama ve Algoritmik Düzeylerde Sosyal Özgünlük.
Şekil 3: Uygulama ve Algoritmik Düzeylerde Sosyal Özgünlük.
(A) Sosyal olarak özelleşmiş bilgi işleme, ayna işleme, kendi kendine işleme veya uygulamada asosyal işleme için seçim davranışının ve / veya beyin tepkisinin varsayılmış modelini gösteren şematik. Bu modeller, “benlik” koşulunun ve / veya “asosyal” bir durumun dahil edilmesinin, belirli bir sürecin sosyal olarak ne kadar özelleşmiş olduğunu ve her durumda görmeyi bekleyeceğimiz beyin veya davranış profili türlerini göstermeye nasıl yardımcı olabileceğini vurgular. (B) Hill ve arkadaşları [71] ile Hampton ve arkadaşlarından [72] “sosyal” ve “sosyal olmayan” algoritma örnekleri: Bu algoritmalar, katılımcıların bir “çalışan” veya “işveren” rolünde başka bir oyuncuyla stratejik olarak etkileşime girdiği bir görevde ayırt edilebilir. Çalışan rolünde, getirilerini en üst düzeye çıkarmak için, katılımcılar denetlendiklerinde çalışmalı ve denetlenmediklerinde çalışmamalıdır. Yazarlar, katılımcıların oyunu nasıl oynadıklarını açıklamak için farklı modelleri karşılaştırdı. Rakibinden (sosyal olmayan) bağımsız olarak ödül sonuçlarını basitçe izleyen basit bir pekiştirmeli öğrenme modeli, oyuncunun stratejisinin rakibin davranışı üzerindeki etkisini hesaba katan bir etki algoritmasının (sosyal) yanı sıra davranışı açıklayamadı. Sosyal örnekte, algoritma, bir temsilcinin duruşmadaki kararını, hem temsilcinin seçim geçmişinin hem de rakibin seçim geçmişinin bir işlevi olarak hesaplar. Bu, rakibin seçim geçmişine bakılmaksızın ortamdaki sonuçların geçmişini basitçe hesaplayan sosyal olmayan bir algoritma ile karşılaştırılır. Pt, rakibin bir eylem seçme olasılığıdır (incelenen veya incelenmeyen). α, öğrenme hızı parametresidir. K, ikinci dereceden inançları tartan ve rakibin parametrelerine (öğrenme oranı, sıcaklık ve kazanç matrisi) yaklaşan bir sabittir. Qt, çalışanın t denemesindeki eylemidir ve qt, işverenin çalışanın çalıştığına dair tahmin ettiği olasılıktır. Sosyal olmayan örnekte, Vt, eylemin seçilip seçilmediğine ve ardından ödülün gelip gelmediğine ilişkin tahmin hatasına (PE: prediction error) dayalı olarak güncellenen eylem değerlerini temsil eder. Tahmin hatası, bir öğrenme oranı (α) ile ağırlıklandırılır. Daha fazla ayrıntı için Hill ve ark. [71] ve Hampton ve ark. [72].
Sosyal olmayan kontrollerle paradigmaları kullanarak ve hem sinir kayıtlarını hem de nedensel yöntemleri kullanarak, bu çalışmalar, uygulama düzeyinde sosyal özgüllük için en açık durumlardan birini sağlar (Şekil 3); ACC’de insanlar, primatlar ve kemirgenler arasında başkalarının acılarına ve ödülüne spesifik olarak yanıt veren hücreler vardır ve ACC hasarı seçici olarak sosyal bilgi işlemeyi bozar.
Farklı Uygulama Ama Aynı Algoritma (?)
İnsanlarda model tabanlı fMRI uygulamasının geliştirilmesi, farklı rakip algoritmaların davranışı açıklayıp açıklayamayacağını ve fonksiyonel anatomi ile eşleştirip eşleştiremeyeceğini test eden birçok çalışma yürütüldü [56, 57, 58]. Pekiştirmeli öğrenme, belki de açık bir algoritmik sürecin en iyi örneğidir ve kendi kendine uygun ve sosyal davranışı anlamak için kapsamlı bir şekilde uygulanmıştır. Pekiştirmeli öğrenme, beklenmedik oluşa (“tahmin hataları”) ve sonuçların değerine dayalı olarak eylemlerin sonuçlarla nasıl ilişkilendirildiğini açıklar, davranışların ödül veya cezalarla nasıl olumlu veya olumsuz olarak nasıl pekiştirildiğini ölçer [18, 57, 58]. İnsanlarda kanıtlar, uygulama için nispeten yüksek düzeyde sosyal özgüllük gösteren aynı bölgenin, ACCg’nin, bunu ortak bir RL algoritması altında yapabileceğini göstermektedir [17, 21, 23, 24, 39, 59, 60]. Bu, uyaranları başkalarıyla ilişkilendirmeyi öğrenmek [23] ile kendi kendine uygun öğrenmeyi karşılaştırmayı, başkalarının tavsiyelerine güvenip güvenmeyeceğinizi [17] ve belirli sosyal tahmin hatalarının (bkz. Sözlük) öğrenmeyi yönlendirdiği yerlerde başkalarına neyi seçeceklerini öğretmeyi [24] içerir. Bu model, pekiştirmeli öğrenme sinyallerini takip etmekle tekrar tekrar ilişkili olan ancak sosyal olarak özelleşmiş bir uygulama olmaksızın ventral striatum gibi diğer beyin alanlarıyla karşılaştırılabilir. Örneğin, birkaç çalışma, gözlem sırasında sosyal öğrenmenin, toplum yanlısı davranışların, özelliklerin anlaşılmasının, güven öğreniminin ve ödüllerle ilgili sosyal olmayan öğrenmenin genellikle bu alanda kodlandığını göstermiştir [17,19, 22, 25, 61, 62]. Ventral striatum tepkileri, hiçbir birey bir sonuçla ilişkilendirilmediğinde tahmin hatalarını bile izliyor gibi görünmektedir [22]. Bu durum, insanlarda, genel-alan öğrenme algoritmasının ventral striatumda uygulanabileceği fikriyle tutarlıdır. Aksine, ACCg’den elde edilen kanıtlar, beyin bölgelerinin sosyal olarak özelleşebileceği, ancak özel bir algoritma uygulamaya ihtiyaç duymayabileceğine işaret eder.
Hücrelerin ve Alanların Ötesinde Sosyal Olarak Özelleşmiş Uygulamalar
Optogenetik, psikofizyolojik etkileşimler, difüzyon tensör görüntüleme ve senkronizasyon ölçümleri gibi metodolojik gelişmeler, uygulamaların yalnızca tek hücrelerde veya beyin alanlarında değil, devrelerde de sosyal olarak özelleşip özelleşmediğini test etmeye olanak tanır. İnsan dışı primatlar ve kemirgenler üzerinde yapılan çeşitli çalışmalar, sosyal olarak özelleşmiş devre uygulamalarına işaret etmektedir. Örneğin, sosyal ödül kodlaması için ACCg’nin sosyal özelleşmesi, bir ACCg-amigdala ağındaki bölgeler arası eşleşme modellerine kadar uzanır [63] ve ACCg ile bazolateral amigdala (BLA) arasındaki projeksiyonlar, klasik korku öğrenmeye kıyasla gözlemsel amaçlıdır [ 52]. Bir BLA → mPFC[1] yolağının aktivasyonu anksiyete benzeri davranışları arttırır ve sosyal etkileşimi azaltırken, aynı yolu engellemek sosyal etkileşimi artırır ve anksiyeteyi azaltır [64]. Bu durum, araştırmanın amigdala düşünüldüğünde sosyal özgüllük eksikliğini öne sürmesine rağmen [11], medial prefrontal alanlarla etkileşime girerken sosyal olarak özelleşmiş devre düzeyinde uygulamalar olabileceğini göstermektedir. İnsanlarda, daha az çalışma, sosyal ve sosyal olmayan bağlanabilirliği doğrudan karşılaştırmıştır. Öte yandan, dikkate değer bir örnek olarak, yalnızca bir başkasının beklenmedik sonucu işlendiğinde mevcut olan ACCg — rostral singulat bağlanabilirliği (ACCg–rostral cingulate connectivity)[2] ve tercihen ortak bir çağrışımsal sinir ağı bulunmaktadır. Doğrudan korku öğrenmeye kıyasla sosyal ortamda temporoparietal bileşke (junction) (TPJ) ile bağlantılıdır [20]. Bunlar, beyin devrelerinde uygulanan sosyal özgüllüğün yalnızca bazı örnekleridir.
Sosyal Olarak Özelleşmiş Algoritmalar
Şimdiye kadar, hücreler, bölgeler ve devrelerin uygulama düzeyinde potansiyel sosyal özgüllüğü tartıştık. Bununla birlikte, sosyal olarak özelleşmiş algoritmalar var mıdır? Bu soru, algoritmaları açıkça tanımlama açısından daha zordur ve türler arasında karmaşık algoritmaların kullanımına ilişkin geniş tartışmalar vardır [4, 66]. Bununla birlikte, zihin teorisi veya zihinselleştirme (mentalizing) [67, 68, 69, 70] gibi bazı sosyal süreçlerin sosyal olarak özelleşmiş olabileceği ve bu nedenle özel algoritmalara dayandığı uzun zamandır tartışılmaktadır. (Figür 3).
Zihin işleme model teorisine yönelik algoritmalar geliştirmeye başlayan birkaç araştırma hattı iki kişilik sosyal alışverişlerdeki davranışı açıklamaya çalışıyor [4, 10, 40]. Bu modellerin çoğu, ekonomik tercihleri veya daha standart pekiştirilmeli öğrenmeyi açıklamak için kullanılan modellerden türetilmiş olsa da, açıkça farklı ve daha karmaşıktır [71, 73] ve sonuçta, bir öznenin neden sosyal etkileşimler dışında bu kadar karmaşıklığa ihtiyaç duyduğu açık değildir. Örneğin, bazı çalışmalar [70, 71, 72], katılımcıların (bir çalışan rolünde) çalışmayı veya kaçmayı seçtikleri ve yaptıklarını inceleyip denetleyemeyen işveren ile etkileşime girdikleri bir işveren veya çalışan rolünü oynamalarını gerektirmiştir (Şekil 3B). Çalışan için, denetlenmediklerinde işten kaçarlarsa ve denetlendiklerinde çalışırlarsa ödüller maksimize edilmiştir. Katılımcıların davranışını en iyi açıklayan algoritma, çalışanın kendi eylemlerinin işverenin davranışı üzerindeki etkisini hesaba katmış ve bu, yalnızca sonuçların geçmişini hesaba katan basit bir pekiştirmeli öğrenmeden daha iyi bir öngörü sunabilmiştir.
Diğer çalışmalar, sosyal olarak özelleşmiş bir düzeneği (saklambaç) sosyal olmayan bir düzenek (kumar) ile doğrudan karşılaştırmış ve katılımcıların zihinselleştiren öznelere karşı daha fazla kazandıklarını göstererek, sosyal etkileşimlerde öğrenirken zihinselleştirmeyi kullanmanın bir katma değerine işaret etmiştir [73]. Bu çalışmanın yazarları ayrıca, insan olmayan primat türlerinin, diğerlerinin kendi eylemlerine vereceği olası tepkilere ilişkin tahminlerini düzenliyormuş gibi davrandıkları yani zihin teorisi algoritmalarının [66] öncü bir biçimini sergilediklerini göstermiştir. Kemirgenlerde, zihin teorisinin varsayılan bir öncüsü çok daha az belirgindir ve bilgimize göre test edilmemiştir. Devaine ve meslektaşlarına göre kemirgenler, sosyal etkileşimde kullanılabilecek çok temel bir algoritmaya ve zihin öncülünün en ilkel teorisine karşılık gelen pekiştirmeli öğrenmeyi sergileyebilirler [66]. Kemirgenlerin bu temel çağrışım sürecini, kendilerini diğerlerinden ayıran çağrışımları öğrenmek için genişletip genişletemeyeceklerini veya zihin teorisine sahip olmak için gerekli olan, kendileri olmayan başka bir hayvanın konseptini taşıyıp taşımayacağını test etmek ilginç olurdu. Kemirgenlerin kendilerine veya ortak kemirgenlere verilen elektrik şoklarının sosyal olarak özelleşmiş modülasyonunu gösteren ve yukarıda bahsedilen gözlemsel öğrenme araştırması göz önüne alındığında, kemirgenlerin bu kapasiteye sahip olması makuldür [50, 51]. İnsanlarda, TPJ’de basitçe kendilik, yakın diğerleri ve uzaktaki diğerleri arasında ilişki kurmanın izini sürerken, ACCg özellikle uyaranlar ve uzaktaki diğerleri arasındaki öğrenme bağlantılarını izlediğine dair kanıtlar vardır [23]. Bu nedenle gelecekteki çalışmalar, kemirgenlerin başka bir hayvan ve bu süreçte yer alan beynin bölümleri hakkında kanıtlanabilir bir şekilde karmaşık bir kavramına sahip olup olmadığını daha fazla araştırabilir.
Zihin teorisinin algoritmaları sosyal olarak özelleşmiş bir şekilde uygulanıyor mu? Çeşitli çalışmalar, TPJ ve dorsomedial prefrontal korteksin (dmPFC) sosyal özgüllük ile zihinselleştirme algoritmalarını işleyebileceğini ileri sürmüştür [4, 10, 40, 67, 74]. Yukarıda bahsedilen iki çalışmada, “etki” algoritması dmPFC ve TPJ’de benzersiz bir şekilde uygulanmıştır [71, 72]. Bununla birlikte, birçok zihin teorisinin — uygulama düzeyinde sosyal özgüllüğü değerlendirmek açısından — bir sınırlaması, genellikle karşılaştırılabilir ‘kendi kendine uygun’ veya ‘sosyal olmayan’ bir koşuldan yoksun olmalarıdır ve bu da özelleşmiş bir uygulamayı sonuçlandırmayı zorlaştırır. (Ek 1) [71, 72, 75]. Dolayısıyla dmPFC veya TPJ’de işlemenin sosyal olarak özelleşmiş mi olduğu, asosyal mi olduğu [23] yoksa kendi kendini izleme ve üstbiliş (bkz. Sözlük) sırasında da devreye giren ortak bir işleme mekanizmasını yansıtıp yansıtmadığı konusunda önemli tartışmalar vardır [76, 77]. “Bilgisayar” aracıları gibi sosyal olmayan kontrol koşullarını kullanan diğer çalışmalar, bir bilgisayara [65, 78, 79, 80] kıyasla “diğer”e daha güçlü yanıtlar göstermiştir (daha fazla tartışma için Ek 1’e bakın). Bu nedenle, zihin teorisinin sosyal özgüllüğünü test etmek için iki gelişme gerekli olacaktır: (i) özgüllüğü uygulama düzeyinde test etmek için uygun sosyal olmayan kontrol koşullarının kullanılması ve (ii) diğer rakiplerin formal algoritmalarının geliştirilmesi algoritmik düzeyde özgüllüğü test etmek için kullanılabilen üstbiliş gibi süreçler.
Marr’ın seviyelerinin bütünleşmesi için ümit vaad eden yeni bir yön; sınıflandırıcılardan, daha model tabanlı olan “temsili benzerlik analizi” (representational similarity analysis RSA) yaklaşımlarına kadar çeşitlenen çok değişkenli tekniklerin kullanılmasıdır. Bu tür yaklaşımlar, bazı bölgelerde faaliyet modellerinin kendilik ve diğerleri arasında farklılaştırılabileceğini göstermek için zaten yararlı olmuştur [81, 82, 83, 84, 85]. Bu, fiziksel acıyı ve sosyal reddi, aynı zamanda ACC’deki [81] benlik ve diğer odaklı değerleri, sağ anterior insulada da [83] başkalarının acısını, tiksinti ve haksız parasal alışveriş dahil olmak üzere diğer olumsuz uyaranları yansıtan modelleri içerir. Farklı benliği ve diğer kalıpları deşifre eden basit sınıflandırıcılar, farklı uygulamayı, yani bir beyin bölgesi içindeki sosyal özgüllüğü yansıtabilir. Bununla birlikte, RSA teknikleri, algoritmik ve uygulama seviyeleri arasındaki boşluğu doldurabilir ve ayrıca algoritmik seviyedeki farklılıkları test edebilir. Bunun nedeni, RSA yaklaşımının, bilgi veya uyaranların nasıl temsil edildiğine, yani bilginin algoritmik olarak nasıl yapılandırıldığına [86] ilişkin test modellerini içermesidir. Farklı algoritmik yapıları tahmin eden rakip modeller üretilebilir ve daha sonra bunlarla en güçlü şekilde ilişkilendirilen beyin bölgeleri ölçülebilir. Yaklaşım, beyin görüntüleme verilerini anlamak için modelleri kullanarak ve tersine, rakip modelleri oluşturmak için beyin görüntüleme verilerini kullanarak algoritmik ve uygulama düzeylerini birbirine bağlayabilir [86].
Bu tür bir yaklaşım, işlev açısından uygulama düzeyi hakkında daha fazla bilgi edinmek için pekiştirmeli öeğrenme gibi belirli bir bilişsel modeli varsayan parametrik model tabanlı görüntüleme yaklaşımları kullanılarak da mümkündür. Bu nedenle modeller, aynı zamanda, yüzler ve evler gibi beyin verileriyle çelişen koşulların standart kategori analizlerine kıyasla algoritmik ve uygulama seviyeleri arasında daha net bir bağlantı sağlar. Gelecekteki araştırmalar, bu çok değişkenli ve model tabanlı teknikleri kullanarak sosyal özgüllük düzeylerini birbirine bağlayabilir. Bu yaklaşımlar aynı zamanda, birbirleriyle olan karşılıklı bağımlılıklarının yanı sıra seviyelerin doğasına da ışık tutabilir ve etkileşime girdikleri kesin yolu vurgulayabilir.
Hedef Değiştirmek Algoritmaları ve Uygulamaları Değiştirir mi?
Marr’ın çerçevesinin en üst düzeyi olan işlemleme düzeyi, bir bilgi işleme sisteminin hedeflerinin önemine işaret eder. Türler arasında, sosyal davranışların hedeflerinin — sosyal motivasyonlar — bir bağlamdan diğerine farklılık gösterebileceği açıktır. Bir dakika rekabet ederiz, sonra da işbirliği yaparız. Bununla birlikte, bir hedefi değiştirmek, diğer iki düzeyde sosyal özgüllüğü değiştirebilir mi? Daha az çalışma bu fikri doğrudan test etse de, sosyal hedefi değiştirmenin gerçekten sinirsel uygulamaları değiştirebileceğine dair ipuçları vardır. Kemirgenlerde, ACC nöronlarının büyük bir kısmı ödüllerin net değerini kodlar — ödülün boyutu, o kaynak için başka biriyle rekabet etmenin maliyetleriyle indirgenir. Ancak, ACC nöronları yalnızca kemirgenlerin rekabet etmesi gerektiğinde bu net değeri kodlar [87]. Maymunlarda, birçok yanal PFC nöronundaki görev sonuç sinyalleri (yani, kazanma veya kaybetme), maymunların ödül için rekabet edip etmediklerine göre belirlenir [88]. İnsanlarda, başkalarıyla işbirliği yapıyor olsak da rekabet ediyor olsak da, dmPFC’nin diğerlerine kıyasla kendimizin performansını izleme kapsamını ayarlar [89].
Sosyal hedeflerin diğer seviyelerde özgüllüğü düzenleme yeteneğini destekleyen bu bulgular, sosyal davranış bozukluklarını ve bunların sosyal benzersizliğini anlamak için potansiyel çıkarımlara sahiptir. Örneğin, hastalar ve kontroller arasındaki nöral uygulamadaki farklılıkları inceleyen grup çalışmalarında, nöral uygulamadaki veya iki grup arasında kullanılan algoritmalardaki farklılıklar algoritmik veya uygulama farklılıkları gibi görünebilir. Aslında gruplar arasında farklı olan ve nöral tepkide değişikliklere neden olan hedeftir. Bunu destekleyen kanıtlar, duygusal el etkileşimlerinin olduğu videoları izlerken psikopati (sosyal bağlama işaret eden bir tür kişilik bozukluğu) olan ve olmayan katılımcıların nöral tepkilerini karşılaştıran bir çalışmadan gelmektedir [90]. Yazarlar, psikopatik suçlulara empati kurmaları ve talimat almamaları talimatı verildiğinde nöral tepkilerdeki grup farklılıklarının önemli ölçüde azaldığını buldular. Bu çalışma, sosyal hedefin değiştirilmesinin uygulamayı nasıl değiştirebileceğini ve sosyal özgüllüğü incelerken gruplar arasındaki motivasyonu eşleştirmenin önemini vurgulamaktadır.
Son Söz
Sosyal özgüllük hakkındaki tartışmalar, on yıllardır sosyal sinirbilim ve psikolojinin merkezinde yer almaktadır. Burada, bu soruları Marr’ın çerçevesi içinde ele almanın, tartışmaları yeniden yapılandırmaya yardımcı olabilecek yeni bir bakış açısı sağladığını ana hatlarıyla açıklıyoruz (bkz. Bu Yazıdaki Önemli Sorular). Bir deneyin Marr’ın hangi seviyelerinde test ettiğini göz önünde bulundurmak ve üç seviyeden birinde kontrol eden ve ayrışan deneyler tasarlamak, türler arasında soruları yeniden formüle etmemize izin verecektir. Pekiştirmeli öğrenmenin işlemleme modelleri ve ekonomik karar verme gibi algoritmik ve işlemleme seviyeleri arasındaki boşluğu doldurmaya yardımcı olabilecek teknikleri ve temsili benzerlik analizi gibi çok değişkenli teknikleri kullanmak, ilerlemek adına önemli olacaktır. Sosyal özgüllüğün nasıl ortaya çıktığı ve türler arasında nelerin korunduğu açık bir sorudur. Nihayetinde, burada özetlenen yaklaşım, uygulamaları, algoritmaları ve işlemlemeleriyle sosyal beyni yeniden tanımlamamıza yardımcı olabilir.
Bu Yazıdaki Önemli Sorular
Algoritmik ve uygulama seviyeleri nasıl etkileşim kurar ve çok değişkenli ve işlemlemesel modelleme yaklaşımları boşluğu kapatmak için kullanılabilir mi?
Algoritma veya uygulamada tür farklılıkları var mı? Primat olmayan hayvanlarda, insanlar ve maymunlar gibi primatlara kıyasla, evrimsel olarak antik subkortikal yapılarda daha fazla özelleşmiş sosyal işlemenin gerçekleştirilmesi mümkündür. İnsan olmayan hayvanlardan elde edilen kanıtlar çoğunlukla uygulama düzeyinde zarif bir sosyal özgüllük bildirmektedir. Farklı algoritmaları test etmek de bilgilendirici olacaktır ve insan olmayanların sosyal olarak özelleşmiş algoritmaları kullanıp kullanamayacağına dair açık bir soru kalır. Bu çaba, sosyal davranışı anlamak için net algoritmalara sahip işlemleme modellerinin kullanımındaki son gelişmelerle kolaylaştırılabilir.
Sosyal olarak özelleşmiş algoritmalar ve uygulamalar doğuştan var ve önceden belirlenmiş mi, yoksa ilişkisel öğrenme yoluyla ortaya çıkabilirler mi?
Bilişsel veya sinirsel bir sürecin sosyal özgüllüğünü oluşturmayı amaçlayan gelecekteki çalışmaları nasıl tasarlamalıyız? Başka bir seviyedeki etkiyi incelerken bir seviyeyi (işlemleme, algoritma veya uygulama) sabit tutmanın önemini ileri sürüyoruz.
Sosyal açıdan benzersiz olan beyin alanları, devreleri veya hücreleri var mı? En güçlü kanıt ACCg için gibi görünüyor. Toplumsal olarak özelleşmiş algoritmalar var mı? En güçlü kanıt, zihin işleme teorisini akla getiriyor.
Beyindeki sosyal ve sosyal olmayan ayrım kategorik mi yoksa sürekli bir ilişki olabilir mi? Bu sorunun yanıtları, algoritmik, işlemlemesel veya uygulama düzeylerinin beyin evriminde sosyal işlevler için nasıl yeniden tasarlandığını daha iyi anlamaya yardımcı olabilir.
Sözlük
Marr’ın seviyeleri
David Marr, bir bilgi işleme sistemi için üç düzeyde açıklama olduğunu öne sürdü. En yüksek seviye, bilgi işleme sisteminin işlemleme veya hedefidir. İkinci seviye, algoritmik veya sistemin uyguladığı kurallardır. Üçüncü seviye, sistemin uygulanması veya fiziksel olarak gerçekleştirilmesidir.
Üstbiliş
İnançlar, arzular ve niyetler gibi zihinsel durumları kendine atfetme yeteneği.
Ayna nöronları
Başlangıçta maymun premotor korteksinde keşfedilen bu nöronların, hem bir eylemi gerçekleştirirken hem de bir eylemi gözlemlerken, nöral olarak benzer şekilde ateşlenmesi söz konusudur.
Optogenetik
Işığa duyarlı iyon kanallarını ifade etmek amacıyla, ışığın genetik olarak değiştirilmiş nöronları kontrol etmek için kullanıldığı biyolojik bir teknik. Bu teknik, kemirgen çalışmalarında yaygın olarak kullanılmaktadır ancak şu anda insanlarda güvenle kullanılması mümkün değildir.
Pekiştirmeli öğrenme
Uyaranlar veya eylemler ile olumlu ve olumsuz sonuçlar arasındaki ilişkileri öğrenmek. Öğrenme, sonucun ne kadar beklenmedik olduğuna bağlıdır.
Sosyal acı
Diğer insanların başına gelen caydırıcı acı verici olaylar.
Sosyal tahmin hataları
Sosyal etkileşimler sırasında ortaya çıkan, “diğerlerini” içeren, beklenen ve gerçek sonuçlar arasındaki farklar.
Sosyal ödül
Sosyal etkileşimden türetilen veya sosyal etkileşim bağlamında elde edilen ödüller.
Zihin teorisi
İnançlar, arzular ve niyetler gibi zihinsel durumları diğer bireylere atfetme yeteneği.
Teşekkür
Bu çalışma, P.L.L.’ye verilen Tıbbi Araştırma Konseyi Bursu, Christ Church Genç Araştırma Bursu ve Christ Church Araştırma Merkezi bursu; M.A.J.A’ya verilen Biyoteknoloji ve Biyolojik Bilimler Araştırma Konseyi (BBSRC) David Phillips Bursu ve Wellcome Trust ISSF bursu; ve S.W.C.C.’ye verilen Ulusal Ruh Sağlığı Enstitüsü bursu tarafından desteklenmiştir. Bütünleştirici Nörogörüntüleme için Wellcome Merkezi, Wellcome Trust’ın temel finansmanı ile desteklenmektedir. Çizimlerimizdeki yardımı için Colin Stanton’a teşekkür ederiz.
Referanslar
Kaynakça
1. B.J. Crespi. The evolution of social behavior in microorganisms Trends Ecol. Evol., 16 (2001), pp. 178–183
2. P. Chen, W. Hong. Neural circuit mechanisms of social behavior Neuron, 98 (2018), pp. 16–30
3. R. Bshary, et al. Social cognition in fishes Trends Cogn. Sci., 18 (2014), pp. 465–471
4. M.K. Wittmann, et al. Neural mechanisms of social cognition in primates Annu. Rev. Neurosci., 41 (2018), pp. 99–118
5. D. Marr. Vision MIT Press (1982)
6. J.W. Krakauer, et al. Neuroscience needs behavior: correcting a reductionist bias Neuron, 93 (2017), pp. 480–490
7. J. Bickle. Marr and reductionism Top. Cogn. Sci., 7 (2015), pp. 299–311
8. C. Lamm, et al. Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain Neuroimage, 54 (2011), pp. 2492–2502
9. Y. Fan, et al. Is there a core neural network in empathy? An fMRI based quantitative meta-analysis Neurosci. Biobehav. Rev., 35 (2011), pp. 903–911
10. C.C. Ruff, E. Fehr. The neurobiology of rewards and values in social decision making Nat. Rev. Neurosci., 15 (2014), pp. 549–562
11. J. Munuera, et al. Shared neural coding for social hierarchy and reward value in primate amygdala Nat. Neurosci., 21 (2018), pp. 415–423
12. M.A. Umilta, et al. I know what you are doing: a neurophysiological study Neuron, 31 (2001), pp. 155–165
13. T.T.-J. Chong, et al. fMRI adaptation reveals mirror neurons in human inferior parietal cortex Curr. Biol., 18 (2008), pp. 1576–1580
14. C. Heyes. What’s social about social learning? J. Comp. Psychol., 126 (2012), pp. 193–202
15. R. Cook, et al. Mirror neurons: from origin to function Behav. Brain Sci., 37 (2014), pp. 177–192
16. C. Catmur, et al. Associative sequence learning: the role of experience in the development of imitation and the mirror system Philos. Trans. R. Soc. B Biol. Sci., 364 (2009), pp. 2369–2380
17. T.E.J. Behrens, et al. Associative learning of social value Nature, 456 (2008), pp. 245–249
18. P.L. Lockwood, M. Klein-Flügge. Computational modelling of social cognition and behaviour — a reinforcement learning primer Soc. Cogn. Affect.
19. A. Olsson, et al.The neural and computational systems of social learning Nat. Rev. Neurosci., 21 (2020), pp. 197–212
20. B. Lindström, et al. A common neural network differentially mediates direct and social fear learning NeuroImage, 167 (2018), pp. 121–129
21. M.A.J. Apps, et al. The anterior cingulate gyrus and social cognition: tracking the motivation of others Neuron, 90 (2016), pp. 692–707
22. P.L. Lockwood, et al. Neurocomputational mechanisms of prosocial learning and links to empathy Proc. Natl. Acad. Sci. U. S. A., 113 (2016), pp. 9763–9768
23. P.L. Lockwood, et al. Neural mechanisms for learning self and other ownership Nat. Commun., 9 (2018), p. 4747
24. M.A.J. Apps, et al. Vicarious reinforcement learning signals when instructing others J. Neurosci., 35 (2015), pp. 2904–2913
25. S. Sul, et al. Spatial gradient in value representation along the medial prefrontal cortex reflects individual differences in prosociality Proc. Natl. Acad. Sci. U. S. A., 112 (2015), pp. 7851–7856
26. A.S. Gabay, M.A.J. Apps. Foraging optimally in social neuroscience: computations and methodological considerations Soc. Cogn. Affect. Neurosci. (2020)
27. S.M. Reader, K.N. Laland. Social intelligence, innovation, and enhanced brain size in primates Proc. Natl. Acad. Sci. U. S. A., 99 (2002), p. 4436
28. R.I.M. Dunbar. Neocortex size as a constraint on group size in primates J. Hum. Evol., 22 (1992), pp. 469–493
29. R.I.M. Dunbar. The social brain hypothesis Evol. Anthropol. Issues News Rev., 6 (1998), pp. 178–190
30. T. Sawaguchi, H. Kudo. Neocortical development and social structure in primates Primates, 31 (1990), pp. 283–289
31. A. Jolly. The evolution of primate behavior: a survey of the primate order traces the progressive development of intelligence as a way of life Am. Sci., 73 (1985), pp. 230–239
32. N.K. Humphrey. The social function of intellect P.P.G. Bateson, R.A. Hinde (Eds.), Growing Points in Ethology, Cambridge University Press (1976), pp. 303–318
33. T. Matsuo, et al. Genetic dissection of pheromone processing reveals main olfactory system-mediated social behaviors in mice Proc. Natl. Acad. Sci. U. S. A., 112 (2015), pp. E311-E320
34. A. Asaba, et al. Sexual attractiveness of male chemicals and vocalizations in mice Front. Neurosci., 8 (2014), p. 231
35. D.J. Horschler, et al. Do non-human primates really represent others’ beliefs? Trends Cogn. Sci. (2020) Published online June 24, 2020.
36. C. Grimm, et al. Shedding light on social reward circuitry: (un)common blueprints in humans and rodents Neuroscientist (2020)
37. I.B.-A. Bartal, et al. Empathy and pro-social behavior in rats Science, 334 (2011), pp. 1427–1430
38. M. Laubach, et al. What, if anything, is rodent prefrontal cortex? eNeuro, 5 (2018), Article e0315–18.2018
39. J. Joiner, et al. Social learning through prediction error in the brain Npj Sci. Learn., 2 (2017), p. 8
40. D. Lee, H. Seo. Neural basis of strategic decision making Trends Neurosci., 39 (2016), pp. 40–48
41. S. van Heukelum, et al. Where is cingulate cortex? A cross-species view Trends Neurosci., 43 (2020), pp. 285–288
42. P.L. Lockwood. The anatomy of empathy: vicarious experience and disorders of social cognition Behav. Brain Res., 311 (2016), pp. 255–266
43. M.A.J. Apps, et al. The role of the midcingulate cortex in monitoring others’ decisions Front. Neurosci., 7 (2013), p. 251
44. A. Burgos-Robles, et al. Conserved features of anterior cingulate networks support observational learning across species Neurosci. Biobehav. Rev., 107 (2019), pp. 215–228
45. S.W.C. Chang, et al. Neuronal reference frames for social decisions in primate frontal cortex Nat. Neurosci., 16 (2013), pp. 243–250
46. R.L. Kendal, et al. Social learning strategies: bridge-building between fields Trends Cogn. Sci., 22 (2018), pp. 651–665
47. P.H. Rudebeck, et al. A role for the macaque anterior cingulate gyrus in social valuation Science, 313 (2006), pp. 1310–1312
48. B. Basile, et al. The anterior cingulate cortex is necessary for forming prosocial preferences from vicarious reinforcement in monkeys PLoS Biol., 18 (2020), Article e3000677
49. P.L. Lockwood, et al. Anterior cingulate cortex: a brain system necessary for learning to reward others? PLoS Biol., 18 (2020), Article e3000735
50. M. Carrillo, et al. Emotional mirror neurons in the rat’s anterior cingulate cortex Curr. Biol., 29 (2019), pp. 1301–1312
51. J. Hernandez-Lallement, et al. Harm to others acts as a negative reinforcer in rats Curr. Biol., 30 (2020), pp. 949–961
52. S.A. Allsop, et al. Corticoamygdala transfer of socially derived information gates observational learning Cell, 173 (2018), pp. 1329–1342
53. O. Yizhar, et al. Neocortical excitation/inhibition balance in information processing and social dysfunction Nature, 477 (2011), pp. 171–178
54. P.H. Rudebeck, et al. Distinct contributions of frontal areas to emotion and social behaviour in the rat Eur. J. Neurosci., 26 (2007), pp. 2315–2326
55. D. Jeon, et al. Observational fear learning involves affective pain system and Cav1.2 Ca2+ channels in ACC Nat. Neurosci., 13 (2010), pp. 482–488
56. Y. Niv, A. Langdon. Reinforcement learning with Marr Curr. Opin. Behav. Sci., 11 (2016), pp. 67–73
57. W. Schultz. Updating dopamine reward signals Curr. Opin. Neurobiol., 23 (2013), pp. 229–238
58. P. Dayan, N.D. Daw. Decision theory, reinforcement learning, and the brain Cogn. Affect. Behav. Neurosci., 8 (2008), pp. 429–453
59. P.L. Lockwood, et al. Encoding of vicarious reward prediction in anterior cingulate cortex and relationship with trait empathy J. Neurosci., 35 (2015), pp. 13720–13727
60. M.R. Hill, et al. Observational learning computations in neurons of the human anterior cingulate cortex Nat. Commun., 7 (2016), p. 12722
61. C.J. Burke, et al. Neural mechanisms of observational learning Proc. Natl. Acad. Sci. U. S. A., 107 (2010), pp. 14431–14436
62. L.M. Hackel, et al. Instrumental learning of traits versus rewards: dissociable neural correlates and effects on choice Nat. Neurosci., 18 (2015), pp. 1233–1235
63. O. Dal Monte, et al. Specialized medial prefrontal–amygdala coordination in other-regarding decision preference Nat. Neurosci., 23 (2020), pp. 565–574
64. A.C. Felix-Ortiz, et al. Bidirectional modulation of anxiety-related and social behaviors by amygdala projections to the medial prefrontal cortex Neuroscience, 321 (2016), pp. 197–209
65. J.H. Balsters, et al. Disrupted prediction errors index social deficits in autism spectrum disorder Brain, 140 (2016), pp. 235–246
66. M. Devaine, et al. Reading wild minds: a computational assay of theory of mind sophistication across seven primate species PLoS Comput. Biol., 13 (2017), Article e1005833
67. J.P. Mitchell. Mentalizing and Marr: an information processing approach to the study of social cognition Brain Res., 1079 (2006), pp. 66–75
68. J. Koster-Hale, R. Saxe. Theory of mind: a neural prediction problem Neuron, 79 (2013), pp. 836–848
69. I.A. Apperly. What is ‘theory of mind’? Concepts, cognitive processes and individual differences: Q J. Exp. Psychol., 65 (2012), pp. 825–839
70. W. Yoshida, et al. Game theory of mind PLoS Comput. Biol., 4 (2008), Article e1000254
71. C.A. Hill, et al. A causal account of the brain network computations underlying strategic social behavior Nat. Neurosci., 20 (2017), pp. 1142–1149
72. A.N. Hampton, et al. Neural correlates of mentalizing-related computations during strategic interactions in humans Proc. Natl. Acad. Sci. U. S. A., 105 (2008), pp. 6741–6746
73. M. Devaine, et al. The social Bayesian brain: does mentalizing make a difference when we learn? Biol., 10 (2014), Article e1003992
74. C.D. Frith, U. Frith. The neural basis of mentalizing Neuron, 50 (2006), pp. 531–534
75. T. Hayashi, et al. Macaques exhibit implicit gaze bias anticipating others’ false-belief-driven actions via medial prefrontal cortex Cell Rep., 30 (2020), pp. 4433–4444
76. C.D. Frith. The role of metacognition in human social interactions Philos. Trans. R. Soc. B Biol. Sci., 367 (2012), pp. 2213–2223
77. C. Heyes, et al. Knowing ourselves together: the cultural origins of metacognition Trends Cogn. Sci., 24 (2020), pp. 349–362
78. J.K. Rilling, et al. The neural correlates of theory of mind within interpersonal interactions Neuroimage, 22 (2004), pp. 1694–1703
79. H.L. Gallagher, C.D. Frith. Functional imaging of ‘theory of mind’ Trends Cogn. Sci., 7 (2003), pp. 77–83
80. M.A.J. Apps, et al. Reinforcement learning signals in the anterior cingulate cortex code for others’ false beliefs NeuroImage, 64 (2013), pp. 1–9
81. C.-W. Woo, et al. Separate neural representations for physical pain and social rejection Nat. Commun., 5 (2014), p. 5380
82. C. Corradi-Dell’Acqua, et al. Cognitive and affective theory of mind share the same local patterns of activity in posterior temporal but not medial prefrontal cortex Soc. Cogn. Affect. Neurosci., 9 (2014), pp. 1175–1184
83. C. Corradi-Dell’Acqua, et al. Cross-modal representations of first-hand and vicarious pain, disgust and fairness in insular and cingulate cortex Nat. Commun., 7 (2016), p. 10904
84. M. Piva, et al. The dorsomedial prefrontal cortex computes task-invariant relative subjective value for self and other eLife, 8 (2019), Article e44939
85. M.A. Thornton, et al. People represent their own mental states more distinctly than those of others Nat. Commun., 10 (2019), p. 2117
86. B.C. Love. The algorithmic level is the bridge between computation and brain Top. Cogn. Sci., 7 (2015), pp. 230–242
87. K.L. Hillman, D.K. Bilkey. Neural encoding of competitive effort in the anterior cingulate cortex Nat. Neurosci., 15 (2012), pp. 1290–1297
88. T. Hosokawa, M. Watanabe. Prefrontal neurons represent winning and losing during competitive video shooting games between monkeys J. Neurosci., 32 (2012), pp. 7662–7671
89. M. Wittmann, et al. Self–other mergence in the frontal cortex during cooperation and competition Neuron, 91 (2016), pp. 482–493
90. H. Meffert, et al. Reduced spontaneous but relatively normal deliberate vicarious representations in psychopathy Brain, 136 (2013), pp. 2550–2562
91. K.R. Popper. The logic of scientific discovery Basic Books (1959)
92. F. Castelli, et al. Movement and mind: a functional imaging study of perception and interpretation of complex intentional movement patterns Neuroimage, 12 (2000), pp. 314–325
93. J. Stanley, et al. Effects of agency on movement interference during observation of a moving dot stimulus J. Exp. Psychol. Hum. Percept. Perform., 33 (2007), pp. 915–92
[1] Medial prefrontal korteks (mPFC)
[2] Bkz.: Rostral ön singulat korteks — The rostral anterior cingulate cortex (rACC): Tüm kortikal alanlardan, ACC’nin duygusal bölümü olan rostral anterior singulat korteks (rACC), duygu ve bilişi bütünleştirmek için ideal olarak limbik ve kortikal yapılar arasında konumlandırılmıştır ve bu nedenle amigdalaya bağlı öğrenmeyi etkilemek için hazırlanmıştır. (Ç.N.)
