Özgün Adı: Neuroscience Needs Evolution
Özet
Sinir sistemi, evrimin bir ürünüdür. Başka bir deyişle, kalıtımın güçlü sınırlamaları altında, uzun bir modifikasyon dizisi boyunca inşa edilmiş ve sürekli olarak yoğun seçilim baskılarına maruz kalmıştır. Bunun bir sonucu olarak, beynin organizasyonu ve işlevleri, beynin kendi tarihsel gelişimi tarafından şekillendirilmiştir.
Çağdaş sistemler nörobiliminde (systems neuroscience) görece daha az dikkate alınan bu olgunun beynin gizemlerini çözmemize yardımcı olabilecek son derece değerli bir araç sunduğunu düşünüyoruz. Nitekim evrimsel tarihin dikkate alınmasının, davranışsal deneyler, anatomik yapı incelemeleri ve sinirsel etkinliğin kaydına dayalı işlevsel çözümlemeler gibi, beyni anlamak için kullanılan diğer entelektüel araçlarla birlikte yer alması gerektiğini savunuyoruz.
Bu giriş yazısında, evrimsel kuramın sinirbilimi hangi açılardan zenginleştirebileceğini somut örnekler üzerinden vurgulayarak evrimin önemine dikkat çekmekteyiz. Özel sayının geri kalanında, söz konusu yaklaşım farklı boyutlarıyla derinleştirilmektedir. Özellikle sinirsel evrimin muhafazakâr (conservative) niteliği, evrimsel tarihimizdeki belirli geçişlerin doğurduğu önemli sonuçlar ve evrimsel bakış açısının, özgül (specific) mekanizmalardan beyin organizasyonunun temel ilkelerine kadar uzanan meseleleri nasıl aydınlattığı tartışılmaktadır.
Bu makale, “Evrimsel kuram merceğinden sistemler sinirbilimi” başlıklı özel sayısından bir kesittir.
Anahtar Kelimeler: psikoloji (psychology), ontoloji (ontology), filogenetik tarih (phylogenetic history), evrimsel nörobilim (evolutionary neuroscience), gelişimsel nörobilim (developmental neuroscience)
1. Giriş
Tüm biyolojik varlıklar gibi, beyin ve sinir sistemi de evrimin ürünüdür. Diğer bir deyişle, doğal seçilim tarafından elenen ve çeşitlilik gösteren biçimlerin ortaya çıkmasına yol açan milyonlarca yıl boyunca süren uzun bir değişim dizisi aracılığıyla yavaş yavaş oluşmuşlardır. Bilim insanlarının neredeyse tamamı bu görüşü kabul etmektedir. Evrim, biyolojik sistemlerin her düzeyde hem temel ilkeleri hem de özgül ayrıntıları için nihai açıklamayı sunar ve biyolojinin “büyük birleştirici kuramı” (grand unifying theory) olarak değerlendirilebilir [1]. Buna karşın, bilim dünyasında fiilen oy birliğiyle kabul edilmesine rağmen, evrimin nörobilim açısından doğurduğu sonuçların yeterince fark edilmediğini düşünüyoruz. Nitekim, özellikle sistemler düzeyindeki (systems-level) nörobilim araştırmalarının önemli bir bölümü, evrimi hesaba katmıyormuş gibi ilerlemektedir. Çoğu durumda bu çalışmalar evrimsel ilkeleri açıkça reddetmez, ancak bu ilkelerin araştırmalarımıza ne ölçüde yön verebileceğini dikkate almaz. Başka bir deyişle, sistemler nörobiliminin büyük bir kısmı elindeki başlıca araçlardan birini kullanmayı ihmal etmektedir.
Evrimin bu ölçüde ihmal edilmesi, kısmen modern sistemler nörobiliminin ortaya çıkmasına zemin hazırlayan çeşitli disiplinlerin etkisinden kaynaklanmaktadır. Özellikle psikoloji, başlangıçta biyolojiye ilişkin sorulardan ayrı olarak insan zihinsel süreçlerinin incelenmesi şeklinde açıkça tasarlanmıştır [2]. Benzer şekilde, modern sistemler nörobilimi üzerinde büyük etkisi olan bilişsel bilim de ilk dönemlerinde zihni, beynin “donanımından” (hardware) ziyade bir “yazılım” (software) meselesi olarak ele almıştır [3,4]. Her ne kadar çağdaş bilişsel nörobilim ve sistemler nörobilimi bu farklı düzeyler arasında bağlantı kurmayı hedeflese de temel kavramlar (örneğin dikkat, çalışma belleği vb.) hâlâ büyük ölçüde psikolojik gelenekler tarafından belirlenen çerçeveler içinde tanımlanmaktadır. Sonuç olarak, hayvan davranışlarının mekanizmaları sıklıkla evrimsel tarih ve biyolojik çeşitlilik göz ardı edilerek insan bilişini açıklamak üzere geliştirilmiş kuramlar üzerinden yorumlanmaktadır.
Nitekim evrimin ihmal edilmesi, nörobilimin büyük ölçüde insan beynini anlamaya yönelmiş olduğu gerçeğini de yansıtmaktadır. Görünürde temel ve sadece meraka dayalı olan araştırmalar dahi, çoğunlukla insan hastalıklarına yönelik tedaviler geliştirme misyonu taşıyan kurumlar tarafından finanse edilmektedir. Temel sistemler nörobilimi yaygın biçimde hayvan modellerinden yararlansa da, bunlar genellikle yalnızca “model” olarak görülmektedir. Bir başka deyişle, bu modeller insan beyninin temsilleri olarak ele alınmakta ve insanlara ne ölçüde genellenebilir oldukları (external validity) ölçüsünde değer kazanmaktadır. Bunun bir sonucu olarak, bu hayvanların beyinlerini birbirinden ayıran özellikler yeterince takdir edilmemekte ve üzerinde yeterince çalışılmamaktadır. Nihayetinde ortaya çıkan tablo ise, uzun bir evrimsel sürecin yalnızca bir basamağı olmak yerine, hedefi sadece bir türe odaklanan bir bilim anlayışıdır.
“Hayvan davranışlarının mekanizmaları sıklıkla evrimsel tarih ve biyolojik çeşitlilik göz ardı edilerek insan bilişini açıklamak üzere geliştirilmiş kuramlar üzerinden yorumlanmaktadır.”
Evrimsel ilkelerin sistemler nörobilimi üzerindeki etkisinin sınırlı kalmasının bir diğer olası nedeni de bu ilkelerin öneminin yeterince bilinmemesidir. Dolayısıyla, bu giriş bölümünün amaçlarından biri, literatürden seçilen çeşitli örneklerle evrimsel düşünme biçiminin nörobilime nasıl rehberlik edebileceğini betimlemektir. Aslında, bu özel sayının tamamının amacı da tam olarak budur.
2. Evrimsel Tarihin Önemi
Doğal seçilim kavramı sistemler nörobiliminde sıklıkla kullanılmaktadır; ancak bu kullanım çoğu zaman, sinirsel sistemlerin belirli performans ölçütleri açısından en uygun (optimality) varsayımını gerekçelendirmekten öteye gitmez. Diğer bir deyişle, evrim doğanın dünyadan kaynaklanan belirli sorunlara en uygun çözümleri bulma süreciymiş gibi ele alınmaktadır. Bu bakış açısında, doğal seçilim yoluyla ulaşılan çözümlerin evrimsel süreçten bağımsız olduğu ve bu süreç dikkate alınmadan da anlaşılabileceği yönünde örtük bir varsayım yatar. Bu şekilde açıkça ifade edildiğinde, söz konusu varsayımın yanlış olduğu bariz biçimde görülmektedir. Evrimin seçici sağkalım (differential survival) dışında bir sorunu çözmeye dair bir hedefi yoktur ve evrim belirli bir problemi tanımlayan herhangi bir ölçüt sunmaz. Bunun yerine, atasal bir sistemin varyasyonlarını üretir. Ardından doğal seçilim yoluyla, kimi zaman daha önce bir sorun olarak görülen durumların üstesinden gelmeyi başaran, işe yarar nitelikler sergileyen varyasyonları kayırır. Evrim ne optimalliğe ne de karmaşıklığa yönelik bir hedef güder, yalnızca çeşitliliğin genişlemesine dair bir yanlılık gösterir. Son derece geniş kaynaklara ve çok uzun zaman ölçeklerine sahip olduğu için evrim nihayetinde hayranlık uyandırıcı sonuçlar ortaya koyabilir. Yine de evrim bizim tanımlayabileceğimiz belirli hesaplamalı (computational) ya da bilişsel problemlere yönelik çözümler bulma süreci değildir.
“Evrimin seçici sağkalım (differential survival) dışında bir sorunu çözmeye dair bir hedefi yoktur ve evrim belirli bir problemi tanımlayan herhangi bir ölçüt sunmaz.”
Buna rağmen, evrimsel sürecin ortaya çıkardığı mekanizmaların bu süreçten bağımsız olduğu ve evrim dikkate alınmadan da anlaşılabileceği hâlâ ileri sürülebilir. Bu argüman ilk bakışta makul görünse de, evrim kuramının bir diğer temel dayanağı olan modifikasyonla türeyiş (descent with modification) gerçeğini göz ardı etmektedir. Buna göre, bir varyasyonun doğal seçilimin oyununa katılabilmesi için öncelikle “mümkün” olması gerekir. Mümkün varyasyonların kapsamı ise atasal sistem tarafından büyük ölçüde sınırlandırılmıştır. Mümkün olan mekanizmaların alanının, yalnızca işlevsel kaygılar veya optimallik temelinde düşünüldüğünden çok daha kısıtlı olmasının birçok nedeni vardır. Özellikle çok hücreli organizmalarda, bu sınırlamanın başlıca kaynaklarından biri gelişimsel süreçtir. Genom, vücut için bir taslak ya da beyin için bir bağlantı haritası (connectome) tanımlamaz; aksine, uzun bir dizi gelişimsel değişim aracılığıyla vücut ve beyni inşa etmenin bir tarifini sunar. Her tarifte olduğu gibi, sonraki aşamalar önceki aşamalara bağımlıdır. Dolayısıyla değişiklikler gelişigüzel bir şekilde sürece dahil edilemez. Gelişimin erken evrelerinde ortaya çıkacak değişikliklerin adaptif olma olasılığı düşüktür. Bunun başlıca nedeni, bu değişikliklerin genellikle sonraki aşamaların işleyişine temel teşkil eden varsayımları ihlal etmesidir. Bu nedenle, türlerde başarılı yeni varyasyonlar genellikle ya gelişimin sonunda yeni aşamalar ekleyen ya mevcut sistemleri kopyalayıp farklı şekilde özelleştiren ya da tüm gelişimsel zincirleri terk eden ve gerileyen değişimlerdir. Sonuç olarak, herhangi bir soy hattı boyunca hayvanların evrimi son derece muhafazakârdır. Bunun bir getirisi olarak modern hayvanların yapıları ve mekanizmaları, büyük ölçüde atalarının yapılarına ve mekanizmalarına bağlıdır ve içinde bulunduğu atasal bağlam dışında anlaşılamaz.
Peki, bu durumun nörobilim için ne gibi sonuçları vardır? Nörobilimciler olarak karmaşıklığı göz korkutan bir kara kutuyla (yani, beyinle) karşı karşıyayız. Ancak elimizde paha biçilmez bir dayanak bulunmaktadır. Bu kara kutu bir anda var olmamış; aksine, zaman içinde, yavaş yavaş, herhangi bir tasarımcının müdahalesi olmaksızın inşa edilmiştir. Buna ek olarak, bu inşa süreci ile gelişimi nedensiz olmayan ve çoğu zaman optimallikten daha fazlasına yönelik katı kısıtlamalar içerisinde gerçekleşmiştir. En önemlisi de, bu sürecin adımlarını makul bir güvenilirlikle tahmin edebilecek yaklaşımlar ve halihazırda onlarca yıldır bunu yapan muazzam bir külliyat mevcuttur. Bu çalışmalar, aksi takdirde anlaşılmaz kalacak olan yapı ve işleve dair soruları yanıtlamamıza yardımcı olabilecek içgörüler sunmaktadır [5]. Bu tür içgörüleri göz ardı etmek ve beynin biyolojik gerçekliğiyle bağdaşmayan geniş, kısıtlamalardan yoksun bir kuram alanını keşfetmeye çalışmak akıllıca görünmemektedir.
Evrimsel tarihin dayattığı katı kısıtlamalara dair ünlü örneklerden biri omurgalıların retina yapısıdır. Her birinci sınıf sinirbilim öğrencisinin bildiği gibi, retina ters bir organizasyona sahiptir. Başka bir deyişle, birincil duyu hücreleri (primary sensory cells) mercekten uzağa yönelmiş durumdadır ve ışığın ışığa duyarlı (photosensitive) reseptörlere ulaşabilmesi için yatay , bipolar, amakrin ve gangliyon hücre katmanlarından geçmesi gerekir. Peki, bunun adaptif nedeni ne olabilir? Filtrelemeden korunmaya kadar pek çok açıklama öne sürülmesine rağmen gerçek yanıt çok daha basittir. Aslında bunun adaptif bir nedeni yoktur. Retinanın bu ters düzeni, sadece gözün evrimsel tarihinin bir sonucudur [6]. Şekil 1a’da gösterildiği gibi, fotoreseptörler başlangıçta sinir plağından dışa doğru yönelmişti. Ancak sinir tüpü (neural tube) oluşurken içe doğru katlandı. Bu durum, neredeyse tamamen saydam olan ve görme işlevini yalnızca günlük ritimleri (circadian rhythms) düzenlemek veya ara sıra geçen gölgelerden kaçmak için kullanan ilkel kordalı (ancient chordate) atalarımız için bir sorun teşkil etmiyordu. Ancak o zamandan beri, fotoreseptörler bu içe dönük konumda sıkışmış durumdadır ve doğal seçilim görsel devrelerin kurulması için gereken gelişimsel sürecin geri kalanını bozmadan onları basitçe ters çevirememiştir. Sinir tüpünün bazı kısımları dışarıya doğru göç ederek yan gözleri oluşturduğunda (Şekil 1b) en az zahmetli yol, reseptör olmayan katmanları saydam bırakmak oldu. Böylece ters retina yapısı kediler, şahinler ve insanlar gibi görsel açıdan etkileyici bazı hayvanlarda bile korunmuştur. Buna karşılık, nörulasyon geçirmeyen kafadanbacaklılarda (cephalopods) retinal reseptörler dışa doğru yönelmiştir. Omurgalı retinası, Dobzhansky’nin ünlü sözünü doğrular niteliktedir: “Biyolojide hiçbir şey, evrim ışığı olmadan anlam kazanmaz.” [5]. Benzer örnekler, nörobilim de dahil olmak üzere biyolojinin tüm alanlarında bolca mevcuttur.
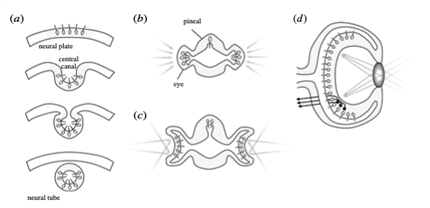
Şekil 1. Omurgalı gözünün evrimsel tarihi [6].
(a) Eumetazoa apikal sinir sistemine ait ışığa duyarlı hücreler (üstte), kordalılarda sinir tüpünün (neural tube) oluşumu sırasında içe doğru katlanmış (altta) ve böylece silli reseptörleri merkezi kanala doğru yönelmiştir.
(b) Sefalatlarda, rostral sinir tüpü yanlara doğru genişleyerek gözleri, dorsal yönde ise pineal bezi oluşturmuştur.
(c) Bunu takiben, lateral gözler içe doğru katlanarak kadeh şeklini almış; böylece duyusal yüzeyin farklı bölümleri dünyanın farklı bölgelerine maruz kalarak uzamsal topografi (spatial topography) yeteneği kazanmıştır.
(d) Omurgalı gözü daha da içe doğru katlanmış ve açıklığın üzerinde bir mercek oluşmuştur. Bu sayede görüntülerin retina yüzeyine odaklanmış bir şekilde yansıtılmasına olanak tanınmıştır.
“Biyolojide hiçbir şey, evrim ışığı olmadan anlam kazanmaz.”
Theodosius Dobzhansky
Nörobilimde tarihin önemine dair bir diğer örnek, belleğin işlevsel nöroanatomisi üzerine yapılan araştırmalar bağlamında karşımıza çıkmaktadır. Geleneksel bellek anlayışı, beyinde tek bir olaysal bellek (episodic memory) sisteminin bulunduğunu ve bunun medial temporal lobda yer aldığını öne sürer [7]. Murray ve arkadaşları [8], bu görüşün mantıklı olsa da primatların evrimsel tarihi hakkında bildiklerimizle tam olarak uyuşmadığını ileri sürmektedir. Nitekim evrimsel değerlendirmelerin geleneksel görüş üzerinde ciddi şüpheler uyandırmaya yeteceğini ileri sürmektedirler. Bunun yerine, primatların öğrenme sistemlerinin ve beynin evrimi hakkındaki bilgilerimizden yararlanarak alternatif bir bakış açısı öne sürmek için kullanılabileceğini savunmaktadırlar.
Aynı araştırmacılar, farklı zamanlarda evrimleşmiş ve farklı işlevler üstlenen yedi ayrı bellek sistemimiz olduğunu öne sürmektedir. Bu sistemler şunlardır: (1) pekiştirme belleği (reinforcement memory), (2) yönelim belleği (navigation memory), (3) yanlı rekabet belleği (biased competition memory), (4) el ile besin arama belleği (manual foraging memory), (5) özellik belleği (feature memory), (6) hedef belleği (goal memory) ve (7) sosyal-öznel bellek (social-subjective memory). Bu bellek sınıflandırması, belleği genel özellikleri üzerinden kategorize eden, kuramsal ilkelere dayalı geleneksel yaklaşımdan farklıdır. Geleneksel yaklaşım, belleği genel özelliklerine göre kategorize ederken, bu sınıflandırma evrimsel anlayışa dayanmaktadır. Nitekim her bellek türü evrimsel tarihte belirli bir döneme karşılık gelir. Örneğin, pekiştirme belleği, hareket kontrolü ile birlikte ilk evrimleşen sistemdir, yönelim belleği ise erken canlıların sadece yerel hareketlerle sınırlı kalmayıp yön bulmaya başlamasıyla daha sonra ortaya çıkmıştır. Evrimsel süreçte son olarak, (7) sosyal-öznel bellek sistemlerimiz insan soy hattında, yani son 5 milyon yıl içinde evrimleşmiştir. Dikkate değer olarak, bu yedi bellek türü ayrı beyin katmanlarına karşılık gelmektedir. Örneğin, (2) navigasyon belleği hipokampal kompleks ile ilişkilendirilirken, (3) yanlı rekabet belleği agranüler prefrontal korteks ile bağlantılıdır. Elbette, bu bellek türlerinin tek bir beyin bölgesiyle tam olarak örtüştüğü söylenemez. Örneğin, sosyal-öznel belleğin granüler prefrontal bölgelerle ilişkili olduğu öne sürülmektedir.
Murray ve arkadaşlarının bellek sistemlerine dair sundukları argümanlar, bellek nörobilimiyle sınırlı kalmayan önemli bir noktaya parmak basmaktadır. Özellikle, evrimsel tarihe dair bilgimiz eksik olsa da bu tarihin nörobilimdeki sorulara yanıt ararken bize yol gösteren değerli bir bilgi kaynağı sunduğunu göstermektedirler. Üstelik bu yardım, retina ve erken görme sistemi gibi çevresel (peripheral) sistemlerle sınırlı değildir. Evrim, üst düzey bilişsel süreçleri (higher-level cognition) bile anlamamıza yardımcı olabilir.
Tıpkı evrimin mikroskobik ölçekteki devreleri ve makroskobik ölçekteki işlevsel anatomiyi anlamamıza yardımcı olabileceği gibi davranışı anlamada da yol gösterici olabileceği görülmektedir [9,10]. Bunun çarpıcı bir örneği, karar verme üzerine yapılan nörobilim ve psikoloji araştırmalarından gelmektedir [11,12]. Kuşkusuz, bu konu nörobilim alanındaki en öne çıkan başlıklardan biridir. Çoğu zaman kararlarımız elimizdeki seçeneklerin değerlerinden etkilenir ve hatta bu değerler tarafından belirlenir. Bu tür seçimleri nasıl yaptığımıza dair tipik yaklaşım ise ekonomi ilkelerine dayanmaktadır. Evrimden ziyade kuramsal ilkelerden türetilen bu geleneksel yaklaşım en basit ve temel karar probleminin eşzamanlı sunulan iki seçenek arasında yapılan bir tercih olduğunu savunur. Karar verme alanının geri kalanı ise bunun bir uzantısı olarak ele alınır. Bu görüşe göre beyin, seçeneklerin öznel faydasını (subjective utility) ortak bir değer birimi (common currency) hesaplar, bu ölçekte daha büyük değere sahip olanı seçer ve ardından bu seçimi bir eylem planına dönüştürür [13-15].
Bununla birlikte, bu görüş çok sayıda kuramsal ve ampirik çalışmanın temelini oluştursa da besin arama kuramının (foraging theory) ilkeleriyle tutarlı değildir [16]. Besin arama kuramı, mikroekonomiden esinlenen ancak biyolojik gerçeklikler ve evrim ilkeleri doğrultusunda şekillenmiş bir davranışsal ekoloji dalıdır [17]. Burada önemli olan husus, doğal koşullarda hayvanların aralarından sadece birini seçebilecekleri iki seçenekle eş zamanlı olarak nadiren karşılaşıyor olmasıdır. Bunun yerine, seçenekler çoğunlukla tek tek ortaya çıkar. Bir seçenek belirdiğinde, karar verici ya onu kabul etmeli ya da reddedip yoluna devam etmelidir. Dolayısıyla besin arama kuramı, ekonomi teorisinin aksine, bir “kabul-ret” kararı temeli üzerine kuruludur.
Kacelnik ve arkadaşları [11], “kabul-ret” öğelerine dayanan zihinsel seçim modellerinin iki seçenekli tercihlere dayalı modellerden çok farklı olduğunu ustalıkla göstermişlerdir. Karar vericiler, görünüşte ikili bir seçim durumunda bile çevrenin zenginliğine dair tahminlerine dayanarak içsel bir eşik değeri belirler ve karşılaştıkları ilk seçeneği bir kabul-ret kararı olarak değerlendirirler. Ardından, ilk seçeneğin değeri hakkındaki bilgiyi eşik değerlerini güncellemek için kullanır ve ikinci seçeneği değerlendirme sürecine geçerler. Böylece birini seçene kadar seçenekleri sırasıyla değerlendirmeyi sürdürürler. Bu ilkeler, yüzeyde anormal gibi görünen bazı davranış örüntülerine hazır bir açıklama sunmakta [18] ve eşzamanlı seçim paradigmalarında dahi seçeneklerin ardışık olarak değerlendirilmesini vurgulayan sinirsel modelleri (neural models) desteklemektedir [19-21]. Bu ilkeler aynı zamanda seçim süreçlerinin sonuçlarını belirlemede sağlarlıkların (affordances) ve bunlar arasındaki rekabetin rolünü öne çıkaran seçim modellerinin başarısını açıklamaya da yardımcı olmaktadır [22]. Bu tür seçim modelleri genellikle daha dayanıklıdır. Bunun yanında laboratuvarda çalışılan, tartışmalı biçimde daha az doğal olan ayrık bağlamlardan ziyade, kesintisiz seçim bağlamlarında davranışı ve sinirsel etkinliği anlamak açısından umut vaat etmektedir [23].
Nitekim, evrimsel ilkelerin yol göstericiliğinin psikolojik kavramların temel organizasyonunu dahi yeniden gözden geçirmemize yol açacak kadar güçlü olduğuna inanıyoruz. Başka bir deyişle, evrimsel bakış açısı beynin biyolojik doğasını göz önünde bulunduran ve felsefe tarihinin ön kabullerinin yükünü taşımayan bir yaklaşımla, beyin hakkında sorduğumuz soruları dahi yeniden gözden geçirmemize yardımcı olabilir [24]. Yukarıda da belirtildiği gibi, sistemler nörobiliminin kavramsal temellerinin büyük bir kısmı psikolojiden gelmektedir. Psikoloji bilimi bellek, dikkat, algı gibi insan zihinsel süreçlerini betimlemekte işlevsel olan kavramlarla şekillendirilmiştir. Sistemler nörobiliminin hedefi de sıklıkla bu süreçleri hayata geçiren sinirsel mekanizmaları bulmak olarak ifade edilir, fakat bu süreçlerin hatalı tanımlanması durumunda bu hedefe ulaşmak mümkün değildir. Örneğin, içgözlem (introspection) algılarımızın “dikkat” adını verdiğimiz bir süzgeçten geçtiğini düşündürse de, birçok çalışma dikkatin beyinde gerçek bir varlığa ya da ayrı bir sisteme karşılık gelip gelmediğini sorgulamaktadır [25-29]. Buna karşılık, evrimsel tarihe bakıldığında, dikkatle ilişkilendirilen bazı olguların (örneğin görsel seçici dikkat; visual selective attention) karar verme süreciyle daha yakından ilişkili olduğu görülmektedir [30]. Bu bakış açısı, geleneksel olarak bu kavramları farklı yöntem ve paradigmalar kullanarak birbirinden bağımsız çalışan araştırma grupları arasında iş birliğini teşvik etmektedir. Dolayısıyla evrim, yalnızca sorularımıza yanıt bulmak için yeni yöntemler ve içgörüler sunmakla kalmaz. Aynı zamanda, en başta hangi soruları sormamız gerektiğini daha iyi tanımlamamıza da yardımcı olur.
3. Evrimsel Kuram Merceğinden Sistemler Nörobilimi
Yukarıdaki örnekler ister işlev ve anatomi arasındaki ilişki, ister davranış hakkındaki düşünme biçimimiz üzerine olsun, evrimsel ilkelerin nörobilim için faydalı içgörüler sunma potansiyelini vurgulamaktadır. Bu tür örnekler iki temel işlev görmektedir. İlk olarak, evrimin nörobilime sunabileceği katkıların genel önemini somutlaştırmaktadır. İkinci olarak ise evrimsel yaklaşımın, ortaya çıkan tamamlayıcı soruları yanıtlamak üzere kullanılabilecek özel örnekler sağladığını göstermektedir. İşte bu nedenle, evrimsel içgörüleri nörobilim problemlerini çözmek için kullanmakla ilgilenen araştırmacıların makalelerini içeren Philosophical Transactions B dergisinin bu özel sayısını hazırladık.
Bu araştırmacılar kendilerini belirli bir grubun üyesi olarak tanımlamadıkları gibi, çalışmalarının da (bu liste kapsamlı olmasa da) mutlaka aynı alt disiplinde toplandığını düşünmemektedirler. Bunun yerine, çok çeşitli alt disiplinlerden gelen bu araştırmacıları birleştiren unsur, evrimi ciddiye alma konusundaki ortak kararlılıklarıdır. Dolayısıyla bu özel sayının amacı, evrimsel tarihin dikkate alınmasının sistemler nörobilimindeki sorulara nasıl ışık tutabileceğini vurgulamaktır. Bir bütün olarak ele alındığında, bu makalelerin hem evrimsel ilkelerin kullanılması lehine güçlü bir argüman sunduğuna hem de bir dizi somut örnek teşkil ettiğine inanıyoruz. Dahası, bu çalışmalardaki özgün fikirler gelecekteki araştırmalar için verimli bir çıkış noktası oluşturabilir. Bu bağlamda, bu sayının geleneksel yaklaşımlara kıyasla beynin biyolojik doğasını daha fazla vurgulayan farklı bir sistemler nörobilimi tarzına örnek teşkil edebileceğini düşünüyoruz.
Omurgalı evrimi dikkat çekici derecede muhafazakârdır ve Shreyas Suryanarayana, Brita Robertson ve Sten Grillner [31] bu muhafazakarlığın ne kadar güçlü olabileceğine dair önemli bir çalışma ortaya koymuştur. Bu araştırmacılar sinirsel kayıt (neural recording), anatomik iz sürme (anatomical tracing) ile elektriksel ve optik uyarım gibi en ileri tekniklerden bazılarını kullanarak elde edilmiş etkileyici bir literatürü özetleyip sentezlemektedir. Bu çalışmalar, memeliler ile yaklaşık yarım milyar yıl önce soy hattı bizden ayrılmış bir tür olan lamprey arasında omurgalı beyninin temel hatlarının büyük ölçüde benzer olduğunu göstermektedir. Bu benzerlikler hareket sistemi kontrolünün (locomotor control) temel beyin sapı ve omurilik mekanizmalarını [32], orta beyindeki yönelme, yaklaşma ve kaçınma tepkilerini [33,34] ve bazal gangliyonlar gibi ön beyin devrelerini [35] kapsamaktadır. Buna ek olarak, palliumda yer alan ve memelilerdeki piriform ve neokortikal sistemlerle homolog olduğu öne sürülen olfaktör [36], somatosensoriyel ve görsel sistemler de bu yapısal sürekliliğin parçasıdır [37]. Tüm bu temel devreler, son ortak atamızda zaten mevcut olup yüz milyonlarca yıldır varlığını sürdürmektedir. Bu durum bu beyin bölgelerine işlev atfeden kuramların makul olabilmesi için bu bölgelerin evrimsel geçmişinin dikkate alınması gerektiğini gösterdiğinden oldukça önemlidir. Elbette bu, söz konusu bölgelerin zaman içinde uyum sağlamadığı ya da işlevlerini değiştirmediği anlamına gelmez. Bununla beraber, bu değişimler yalnızca tarihsel kısıtlamalar çerçevesinde gerçekleşmiş de olabilir.
“Tüm bu temel devreler, son ortak atamızda zaten mevcut olup yüz milyonlarca yıldır varlığını sürdürmektedir. Bu durum bu beyin bölgelerine işlev atfeden kuramların makul olabilmesi için bu bölgelerin evrimsel geçmişinin dikkate alınması gerektiğini gösterdiğinden oldukça önemlidir.”
Daha da derin homolojiler Thurston Lacalli tarafından ele alınmaktadır. Lacalli’nin çalışması [38], ataları bizim soy hattımızdan 600 milyon yıldan fazla bir süre önce ayrılmış [39] ve bu sürenin büyük bölümünde filtreyle beslenme (filter-feeding) nişinde kalmış bir omurgasız kordalı olan amfiyoksus üzerine yapılan araştırmaları özetlemektedir [40]. Amfiyoksus, omurgalıların hipotalamik ve hareket kontrol sistemlerinin birçoğuna ve bazı görsel yolların homologlarını barındırmaktadır. Fakat, amfiyoksuslar karmaşık duyu organlarından yoksundur ve yalnızca belirsiz bir telensefalon öncülüne sahiptir. Lacalli, sınırlı beyin kapasitesi ve davranış repertuarına rağmen amfiyoksusun hareketli hayvanlarda duyusal bilincin (sensory consciousness) ilk kez nasıl ortaya çıktığı gibi geniş kavramsal ve felsefi sorulara bir pencere açabileceğini öne sürmektedir. Bu çalışma, her ne kadar fareler veya maymunlar üzerinde çalışan nörobilimcilerin gündelik sorunlarından uzak gibi görünse de, son derece değerli bir karşılaştırma noktası sunmaktadır. Bunun sebebi, uzun evrimsel zaman ölçeklerinde hangi özelliklerin korunma eğiliminde olduğunu ve hangilerinin olmadığını göstermesidir. Nitekim biz, özellikle insanlardan filogenetik olarak uzak türler olmak üzere, çok çeşitli türlere ve onların sinir sistemlerine aşina olmanın görünüşte sadece insan veya primat beyniyle ilgilenen aktif nörobilimciler için bile kıymetli bir kaynak olduğuna inanıyoruz [41-43].
4. Evrimsel Geçmişimizdeki Büyük Geçişlerin Önemi
Evrim yavaş gerçekleşebilir, ancak etkileri son derece derin olabilir. Nesiller boyunca buna eşlik eden adaptasyonlarla birlikte büyük ölçekli dönüşümler meydana getirebilir. Bu özel sayıda, evrimin muhafazakâr doğasını vurgulayan çalışmaların yanında, insan soy hattı boyunca gerçekleşen ve bizi bugün olduğumuz hale getiren bazı büyük evrimsel geçişleri ele alan makalelere de yer verilmiştir. Bu geçişlerin en çarpıcı örneklerinden biri sucul ortamdan karasal ortama geçiştir. Malcolm MacIver ve Barbara Finlay [44] bu geçişin duyu sistemleri, özellikle de görme duyusu üzerindeki etkilerini tartışmaktadır. Işığın suda ve havada kırınım özelliklerindeki farklar nedeniyle, atalarımız karaya çıktıklarında algılanan görsel dünyanın hacmi dramatik biçimde genişlemiş ve duyusal açıdan yaklaşık bir milyon kat artış meydana gelmiştir [45]. Bu genişleme, yönelimin yanı sıra karar verme ve planlama açısından da çok daha fazla fırsat sunmuştur. Öte yandan, çok eklemli uzuvlara duyulan ihtiyaç ve bu uzuvların hareket ve duruşunu kontrol edecek sinirsel devrelerin gerekliliği gibi yeni zorlukları da beraberinde getirmiştir. Tüm bu süreçler, sinir sisteminde büyük bir genişleme ve çeşitlenmeye yol açarak biz dahil günümüzde yaşayan hayvanlarda gözlemlediğimiz özgül nöral yenilikleri ortaya çıkarmıştır.
Sudan karaya geçişin diğer sonuçları, Lucia Jacobs tarafından ele alınmaktadır. Jacobs’ın makalesinde hava solunumunun karasal tetrapodlarda hipokampusun evrimi için nasıl bir zemin hazırladığını ortaya konulmaktadır [46]. Jacobs’ın öne sürdüğü kokuyla yön bulma hipotezi (olfactory navigation hypothesis) [47], koku alma duyusunun yalnızca kokuların tanımlanmasına hizmet etmediğini, temelinde kokuların uzamsal yönelim (spatial navigation) amacıyla kullanıldığını ileri sürmektedir. Atalarımız karaya çıktıklarında, koku örneklemesi solunumla bağlantılı hale gelmiştir. Jacobs, bu durumun hipokampal teta ritimlerini, bu ritimlerin kat edilen mesafeyi takip etmek için nasıl kullanılabileceğini ve nihayetinde memelilere özgü belleğin iskeletini nasıl oluşturduğunu açıklayabileceğini ileri sürmektedir.
Bir diğer önemli geçiş, memelilerin gececil (nocturnal) yaşama çekilmesiyle başlamış ve yaklaşık 200 milyon yıl sonra bazı primat türlerinin yeniden gündüzcül (diurnal) yaşama dönmesiyle devam etmiştir. Bu süreç, Jon Kaas, Hui-Xin Qi ve Iwona Stepniewska tarafından kaleme alınan ve görsel sistemdeki ilgili değişikliklere odaklanan bir çalışmada ayrıntılı biçimde ele alınmaktadır [48].
Özellikle, diğer memelilerden farklı olarak primatlar, henüz gececil oldukları evrede bile iyi bir görme yetisi geliştirmişlerdir. Bu durum, büyük ve öne bakan gözlere sahip olmalarıyla ve neokortekse giden görsel projeksiyonların dengesindeki bir değişimle mümkün olmuştur. Bu yeniden yapılanma, superior colliculus üzerinden geçen yolların önemini azaltırken, daha doğrudan olan retino-genikülo-striat yolunun genişlemesine yol açmıştır. Bunu takiben, görsel işlemenin dorsal akışı parietal ve premotor bölgelerde yer alan çok çeşitli eyleme özgü alanlara doğru genişlemiştir.
Paul Cisek’in çalışması, kordat filtreyle beslenen (chordate filter feeders) atalarımızdan hareketli sucul omurgalılara, karasal tetrapodlara, gececil memelilere ve gündüzcü primatlara kadar uzanan türağacını takip ederek bu geçişlerin birçoğunu özetlemektedir [49]. Cisek, bu sürece bağlı sinirsel yenilikleri en tepesinde primat bilişinin yer aldığı hiyerarşik bir yapı içinde üst üste eklenen yeni devreler olarak kavramsallaştırmamıştır. Bunun yerine, bu yenilikleri giderek daha alt kontrol sistemlerine bölünen genel bir geri bildirim kontrol devresinin kademeli bir uzaması olarak tanımlamaktadır. Bu çerçevede, kontrol hiyerarşisinin en üst düzeyi, davranışsal durumun düzenlenmesine yönelik en eski “hipotalamik” kontrol biçimini temsil etmektedir. Soyut planlama gibi yeni alt bölümler ise kontrolün hem daha çok dış dünyaya hem de daha soyut etkileşimlere doğru genişlemesini sağlayan adaptasyonlar olarak ortaya çıkmıştır. Cisek’e göre ortaya çıkan bu mimari, türe özgü belirli eylemleri yönlendirmeye adanmış paralel kontrol sistemlerinden oluşan atalardan kalma bir organizasyonu korumaktadır. Bu sistemler arasındaki seçim bazal gangliyonlar tarafından düzenlenir. Buna karşılık, seçilen sistem içindeki özgül eylemlerin belirlenmesi her bir kortikal harita içinde gerçekleşen rekabet süreçleriyle sağlanmaktadır.
Giovanni Pezzulo, Thomas Parr ve Karl Friston da tüm bu görüşlere benzer noktaları yinelemektedir. Bu araştırmacılar sinir sisteminin temel organizasyonu olarak geri bildirim kontrolünü ön plana çıkarmakta, ancak bu yaklaşımı öngörüsel işleme (predictive processing) ile genişletmektedirler [50]. Özellikle, öngörüsel işlemlemenin son dönemde ortaya çıkmış bir evrimsel yenilik olmadığını, aksine omurgalıların sinirsel organizasyonunun temel bir ilkesi olduğunu vurgularlar. Bu ilke, alostatik kontrol düzeyinden hem uzamsal hiyerarşi hem de zaman ölçeği açısından genişleyen çoklu duyu-motor döngülere (multiple sensorimotor loops) kadar geliştirilmiştir. Bu bakış açısına göre bilişsel yetiler, eski bir duyu-motor kontrol sisteminin üzerine eklenen yeni bir sistem olarak ortaya çıkmaz. Aksine, mevcut kontrol sisteminin bir bölümünün giderek daha soyut ve uzun vadeli kontrole doğru özelleşmesiyle bir uzantı olarak ortaya çıkar.
David Leopold ve Bruno Averbeck ise hem önceki yaklaşımlarla uyumlu hem de farklı bir bakış açısı sunmaktadır. Söz konusu araştırmacılar omurgalı beyninin kendi kendini nasıl eğittiğini ele almakta ve bu süreci öz-eğitim (self-tuition) olarak adlandırmaktadırlar [51]. Bu yaklaşıma göre hipotalamik sistemler, telensefalik sistemleri modüle ederek onları beslenme, eş arama ve tehditlerden kaçma gibi temel işlevlerin yanı sıra dış çevrede yön bulma ve navigasyon için gereken bilgi türlerini öğrenmeye yöneltir. Ayrıca primat beyninin karmaşıklığının büyük ölçüde bu tür etkileşimlerin karmaşıklığını yansıttığını ileri sürer. Stuart Wilson ve Tony Prescott ise benzer konulara dair daha genel bir kuramsal çerçeve sunmaktadır. Bu çerçevede, farklı zaman ölçeklerinde (temporal scales) işleyen katmanlı kontrol mimarilerinin karmaşık davranışlar üretmek için nasıl koordine olabileceğini matematiksel bir modelle tanımlamaktadırlar [52]. Yavaş işleyen süreçlerin hızlı olanlar üzerinde kısıtlayıcı rol oynayabileceği yaygın olarak kabul edilirken, bu araştırmacılar bunun tam tersinin de geçerli olabileceğini göstermektedir. Bunun sonucunda katı bir hiyerarşinin olmadığı, ancak farklı seviyelerin birbirini karşılıklı olarak kısıtladığı (mutually constrain) bir kontrol mimarisi ortaya çıkmaktadır.
5. Kendimizi Anlamada Evrimin Önemi
Evrimsel tarihimize dair bu ve benzeri içgörüler, primat beyninin kendine has özelliklerini anlamada önemli ipuçları sunmaktadır. Buradaki ana temayı bedenleşme (embodiment) kavramı oluşturur. Bu bağlamda bedensellik beynin işlevlerinin çoğunun nihayetinde dünyayla olan etkileşimlerimizi kontrol etmeye yönelik olduğu önerisidir. Elbette bunun aksi mümkün olamazdı. Özellikle doğal seçilim perspektifinden bakıldığında, varoluşu pasif biçimde düşünme yeteneğimiz ne kadar karmaşık olursa olsun, hayatta kalmayı etkileyen bir davranışsal sonuca dönüşmediği sürece önemi yoktur. Thomas Huxley’in dediği gibi, “Yaşamın asıl amacı bilgi değil, eylemdir.” Bu yaklaşım, insan ve diğer hayvanların en soyut bilişsel yetilerini bile etkileşim bağlamında değerlendirmemizi, dolayısıyla bunların sinirsel mekanizmalarını da bu çerçevede düşünmemizi gerektirir. Örneğin, Justin Fine ve Benjamin Hayden, uzun süredir soyut bilişin merkezi olarak kabul edilen antropoid primatların prefrontal korteksinin hareket kontrolüne yönelik premotor devrelerin bir uzantısı olarak yeniden yorumlanabileceğini öne sürmektedirler [53]. Özellikle frontal lobun lateral ve medial kısımlarını kapsayan anatomik ve fizyolojik gradyanların sürekliliğine ve hareket veya soyut hedeflerin seçimini sağlayan mekanizmaların hesaplama benzerliğine dikkat çekmektedirler. Sonuç olarak, söz konusu bölgelerin sadece ekonomik değişkenleri temsil etmekle kalmayıp, temelinde motor hedeflerin seçilmesini sağlayan hiyerarşik bir organizasyonun parçası olarak çok daha geniş işlevler kapsamında yeniden değerlendirilmesi gerektiğini savunmaktadırlar. Başlıca uzun süredir değer biçme ve kıyaslama ile ilişkilendirilen orbitofrontal korteks (OFC) üzerinde durmaktadırlar. Bu bölgenin eylemleri yönlendiren prefrontal hiyerarşinin başlangıç noktası olarak ekonomik seçim işlevlerinden daha kapsayıcı bir bakışla anlaşılabileceğini göstermektedirler (ayrıca bkz. [54]).
“Yaşamın asıl amacı bilgi değil, eylemdir.”
Thomas Huxley
Benzer bir yaklaşımla Louise Barrett, Peter Henzi ve Robert Barton primatların sosyal davranışlarını anlamanın en iyi yolunun bu davranışları etkileşimsel çerçevede ele almak olduğunu önermektedir. Böylece eylem kontrolü ve seçimine dair işlevsel mimari, daha soyut etkileşim alanlarına doğru genişletilmektedir [55]. Örneğin, aynı araştırmacılar sosyal koordinasyonun bireyin başkalarına göre mekândaki konumunu gelişmiş bir biçimde kontrol etmesini gerektirdiğini belirtmektedir. Bunun daha eski dönemlerden kalma, sağlarlık temelli eylem yönlendirme mekanizmalarının bir uzantısı olarak evrimleşmiş olabileceğini tartışmaktadır. Bu tür bedenleşmiş gereksinimlerin primat sosyal bilişinin evrimleştiği çerçeveyi oluşturduğunu ve bu çerçevenin soyut bilgiyi ve diğer zihinlerin meta-temsilini vurgulayan geleneksel yaklaşımlardan daha iyi bir zemin sunduğunu öne sürmektedir.
Eski soruları yeni çerçevelere oturtan bu tutum daha da ileri götürülebilir. Luiz Pessoa, Loreta Medina ve Ester Desfilis’in önerdiği üzere bu yaklaşım insan psikolojisinin birçok yönünü yeniden değerlendirmek için kullanılabilir [56]. Bu araştırmacılar, zihinsel olguları tartışırken kullanılan temel teorik ayrımların (örneğin dikkat, duygu vb.) çoğunun bilim öncesinde ortaya konan fikirlerden kaynaklandığını ve beyne bakıldığında bulduklarımızla her zaman örtüşmediğine dikkat çekmektedir. Örneğin, algı kavramının beynin birtakım işlevlerini tanımlamak için pratik bir terim olduğunu ileri sürerken, biliş ve eylem gibi süreçlerden belirgin biçimde ayrılmadığını savunurlar. Başka bir deyişle, bilişsel psikolojide kullanılan kategoriler beynin doğal yapısına uygun düşmemektedir. Bundan dolayı farklı araştırmacılara [24,57] paralel olarak, beynin işlevsel mimarisini tanımlamak için geleneksel zihinsel kategorilerden uzaklaşmayı savunmaktadırlar. Ayrıca, beynin uzun evrimsel geçmişi boyunca evrimini şekillendiren karmaşık davranışların düzenlenmesine odaklanmayı önermektedirler.
Joseph LeDoux’un makalesi, bilinç ve özellikle de duygusal durumların deneyimlenmesi bağlamında benzer bir noktaya değinmektedir [58]. LeDoux, bilinci tek bir olgu ve tek bir açıklama ile ele almamak gerektiğini ileri sürmektedir. Ona göre, bilinç aslında evrimsel geçmişlerine göre birbirlerinden ayrışan bir dizi olguyu tanımlamak için kullanılan öncül bir terimdir. Özellikle, tehdit edici uyaranlara verilen birincil tepkinin birçok hayvanda görülen ilkel bir özellik olduğu apaçıktır. Halbuki, korkunun öznel deneyimi bundan tamamen farklıdır ve muhtemelen soy hattımıza çok daha yakın bir dönemde ortaya çıkmıştır. LeDoux bu durumu korku olgusunun ötesine geneller. İnsan duygularını niteleyen bilinçli hislerin yaşamımızda karşılaştığımız önemli durumların bilişsel yorumlarından (cognitive interpretations) kaynaklandığını düşünmektedir.
“LeDoux (…) insan duygularını niteleyen bilinçli hislerin yaşamımızda karşılaştığımız önemli durumların bilişsel yorumlarından (cognitive interpretations) kaynaklandığını düşünmektedir.”
Bu sayıdaki birçok makalede tartışılan kuramsal perspektif ve önerilere ek olarak, bazı çalışmalar belirli verileri ve önemli metodolojik meseleleri de ele almaktadır. Örneğin, Margaret Bryer ve arkadaşlarının makalesi, belirli bir davranışsal yetinin farklı türler arasında nasıl dağıldığını ve bu yetinin tarihsel süreci hakkında neler söylediğini filogenetik analizler aracılığıyla göstermektedir [59]. Aynı araştırmacılar, sayısal niceliklere duyarlılık örneğini ele alarak, türlerin sayısal biliş (numerical cognition) becerilerinin sistematik olarak farklılık gösterdiğini ortaya koymaktadır. Daha da dikkat çekici olanı ise bu fikri sonrasında genişletmeleri ve sayısal biliş yeteneklerini beynin biçimsel özelliklerinden olan nöron yoğunluğu ile ilişkilendirmeleridir. Bu çalışma, kapsadığı geniş tür perspektifiyle alışılmışın dışında kabul edilir. Özellikle 48 farklı türün karşılaştırılması sayesinde, tek veya birkaç türe odaklanan dar odaklı çalışmaların aksine daha sağlam sonuçlar çıkarma olanağı sunmaktadır. Çok sayıda türün dahil edilmiş olması, hakkında çıkarım yapmanın son derece zor olduğu bilinen bilişin evrimi hakkında güvenilir sonuçlara ulaşılmasını sağlamaktadır.
Bununla birlikte, türler arasında davranışsal yetilerin karşılaştırılması başlı başına zordur. İki farklı tür, verilen bir görevi benzer davranışlar üreten farklı mekanizmalar aracılığıyla çözebilir. Bunun tersi olarak benzer iki mekanizma, bir tür için görev sunumunun doğal olmaması nedeniyle (örneğin, bir kemirgen ile bir primata sunulan görsel ayırt etme görevi) farklı davranışlar sergileyebilir. A. David Redish ve arkadaşları, türler arasında karşılaştırma yapmak için her bir türün gerçekleştirdiği hesaplamaları dikkate alan bir yöntem önermektedir [60]. Yani, bazı yöntemler bir görevin bir bağlamdan diğerine veya bir türden diğerine olan geçerliliğini belirler. Bu araştırmacılar, benzer şekilde davranışsal süreçlerin bir hesaplamalı algoritma olarak işlevselleştirmenin (operationalize) mümkün olduğunu ve bunun kendine has bir geçerliliği olduğunu öne sürmektedir. Bilginin entegrasyonu sonucunda bir eşik değerine ulaşılması süreci buna bir örnek teşkil edebilir. Bu tür bir hesaplamasal geçerlilik (computational validity) kemirgenler üzerinde yapılan çalışmalardan elde edilen bulguların insanlarda ruhsal bozuklukların etkili tedavilerine hangi ölçüde aktarılabileceğini belirlemek gibi somut amaçlar için kullanılabilir. Redish ve arkadaşları, bu geçerlilik türünün dış geçerlilik ve görünüş geçerliliği (face validity) gibi diğer geçerlilik türlerinin yanında yer alabileceğini öne sürmektedir. Bu sayede, model organizmalar biyolojik olarak oldukça farklı olsalar bile çıkarımlar yapmamıza olanak sağlayabilmektedir. Okuyucu bu tema sayısında ağırlıklı olarak insanları ortaya çıkaran soy hattıyla ilişkili araştırma sorularına odaklandığımızı ve böcekler, yumuşakçalar ve diğer protostomlara özgü yeniliklere dair çalışmalara yer vermediğimizi fark edebilir. Bunun sebebi, bu hayvanlara uzanan soy hattının bizimkinden çok uzun zaman önce ayrılmış olmasıdır. Bu kopukluk nedeniyle onların evrimsel öyküleri kendine has dönemeç ve yönelimlerle bizimkinden büyük ölçüde farklılaşmıştır. Bunların her biri başlı başına ikinci bir özel sayıyı hak edecek niteliktedir. Ayrıca, kuşlar gibi yaşam tarzlarını radikal bir şekilde değiştiren türlerin aksine kertenkeleler gibi soy hattımızdan ayrıldıktan sonra benzer bir nişte kalan hayvanlara daha fazla odaklandık. Bu tercih, kapsamı ve uzunluğu sınırlamaya yönelik bir stratejiden ibarettir ve bu tür çalışmaların sunduğu değerli içgörüleri hiçbir şekilde küçümseme amacı taşımamaktadır. Nitekim, kuşlar üzerine yapılan çalışmalar, Bryer ve arkadaşlarının yakınsak evrime (convergent evolution) dair son derece güçlü bir örnek sundukları makalenin merkezinde yer almaktadır [59].
6. Sonuç
Herhangi bir nörobilim lisansüstü programı alanın entelektüel temellerini oluşturan bir dizi ders içerir. Bunlar arasında sıklıkla nöroanatomi, moleküler biyoloji, gelişim ve fizyoloji gibi dersler yer alır. Bu temel derslerin önemini yadsımamakla birlikte, evrim ilkelerinin de en az bunlar kadar merkezi bir öneme sahip olduğunu savunuyoruz. Nitekim, yukarıda ve bu özel sayının geri kalanında sunulan örneklerin, sistem nörobilimindeki hemen her tür akıl yürütmenin evrimsel tarihin dikkate alınmasını gerektirdiğine dair güçlü bir gerekçe sunduğunu umuyoruz. Örneğin, öğrenciler yalnızca doğal seçilim kavramını öğrenmekle yetinmemelidir. Bunun yanında, gelişimsel değişimlerin önemini, beyin tiplerindeki geniş çeşitliliği, farklı beyinlerin ortak problemleri çözmek için kullandıkları farklı yolları ve anlamaya çalıştıkları beyinlerin gerçek evrimsel tarihine de hâkim olmalıdırlar.
Sonuç olarak, evrimsel tarihin nörobilim için yeterince merkezi bir öneme sahip olduğunu ve derin biyolojik geçmişe dair eksik bilgimizin beyni anlamlandırma becerimizi sınırlayan bir etken oluşturduğuna inanıyoruz. Bu nedenle, evrimsel tarihe yönelik temel araştırmaların nörobilimin hedeflerini ilerletmek açısından kritik olduğu kanaatindeyiz. Bu tür araştırmaların sağladığı içgörüler anatomi, fizyoloji ve davranış alanlarından elde edilen bilgilerle birbirini tamamlamaktadır. Bundan dolayı, beynin anlaşılmasına yönelik çabalarımızda başvurduğumuz temel bilgi birikiminin ayrılmaz bir parçasını oluşturmalıdır.
Kaynakça
[1] Dennett DC. 1996 Darwin’s dangerous idea: evolution and the meaning of life. New York, NY: Simon & Schuster.
[2] Hothersall D. 2003 History of psychology, 4th edn. New York, NY: McGraw-Hill.
[3] Block N. 1995 The mind as the software of the brain. In Thinking: an invitation to cognitive science (eds EE Smith, DN Osherson), pp. 377–425. Cambridge, MA: MIT Press.
[4] Gardner H. 1987 The mind’s new science: a history of the cognitive revolution. New York, NY: Basic books.
[5] Dobzhansky T 1973 Nothing in biology makes sense except in the light of evolution, Am. Biol. Teacher 35, 125–129. JSTOR 4444260; reprinted in Zetterberg, JP (ed.) 1983 Evolution versus creationism. Phoenix, AZ: ORYX Press. (doi:10.2307/4444260)
[6] Lamb TD. 2013 Evolution of phototransduction, vertebrate photoreceptors and retina. Prog. Retin. Eye Res. 36,52–119. (doi:10.1016/j.preteyeres.2013.06.001)
[7] Squire LR, Wixted JT, Clark RE. 2007 Recognition memory and the medial temporal lobe: a new perspective. Nat. Rev. Neurosci. 8, 872–883. (doi:10.1038/nrn2154)
[8] Murray EA, Wise SP, Graham KS. 2017 The evolution of memory systems: ancestors, anatomy, and adaptations. Oxford, UK: Oxford University Press.
[9] Gomez-Marin A, Ghazanfar AA. 2019 The life of behavior. Neuron 104, 25–36. (doi:10.1016/j. neuron.2019.09.017)
[10] Krakauer JW, Ghazanfar AA, Gomez-Marin A, MacIver MA, Poeppel D. 2017 Neuroscience needs behavior: correcting a reductionist bias. Neuron 93, 480–490. (doi:10.1016/j.neuron.2016.12.041)
[11] Kacelnik A, Vasconcelos M, Monteiro T, Aw J. 2011 Darwin’s ‘tug-of-war’ vs. starlings’ ‘horse-racing’: how adaptations for sequential encounters drive simultaneous choice. Behav. Ecol. Sociobiol. 65, 547–558. (doi:10.1007/s00265-010-1101-2)
[12] Cisek P, Pastor-Bernier A. 2014 On the challenges and mechanisms of embodied decisions. Phil. Trans. R. Soc. B 369, 20130479. (doi:10.1098/rstb. 2013.0479)
[13] Padoa-Schioppa C. 2011 Neurobiology of economic choice: a good-based model. Annu. Rev. Neurosci. 34, 333–359. (doi:10.1146/annurev-neuro-061010-113648)
[14] McNamee D, Rangel A, O’Doherty JP. 2013 Category-dependent and category-independent goal-value codes in human ventromedial prefrontal cortex. Nat. Neurosci. 16, 479–485. (doi:10.1038/nn.3337)
[15] Levy DJ, Glimcher PW. 2012 The root of all value: a neural common currency for choice. Curr. Opin. Neurobiol. 22, 1027–1038. (doi:10.1016/j.conb.2012.06.001)
[16] Hayden BY. 2018 Economic choice: the foraging perspective. Curr. Opin. Behav. Sci. 24, 1–6. (doi:10. 1016/j.cobeha.2017.12.002)
[17] Stephens DW, Krebs JR. 1985 Foraging theory. Princeton, NJ: Princeton University Press.
[18] Stephens DW, Anderson D. 2001 The adaptive value of preference for immediacy: when shortsighted rules have farsighted consequences. Behav. Ecol. 12, 330–339. (doi:10.1093/beheco/12.3.330)
[19] Krajbich I, Armel C, Rangel A. 2010 Visual fixations and the computation and comparison of value in simple choice. Nat. Neurosci. 13, 1292–1298. (doi:10.1038/nn.2635)
[20] Rich EL, Wallis JD. 2016 Decoding subjective decisions from orbitofrontal cortex. Nat. Neurosci. 19, 973–980. (doi:10.1038/nn.4320)
[21] Hayden BY, Moreno Bote R. 2018 A neuronal theory of sequential economic choice. Brain Neurosci. Adv. 2, 1–15. (doi:10.1177/2398212818766675)
[22] Cisek P, Kalaska JF. 2010 Neural mechanisms for interacting with a world full of action choices. Annu. Rev. Neurosci. 33, 269–298. (doi:10.1146/annurev.neuro.051508.135409)
[23] Yoo SBM, Hayden BY, Pearson JM. 2021 Continuous decisions. Phil. Trans. R. Soc. B 376, 20190664. (doi:10.1098/rstb.2019.0664)
[24] Cisek P. 2019 Resynthesizing behavior through phylogenetic refinement. Attent. Percept. Psychophys. 81, 2265–2287. (doi:10.3758/s13414-019-01760-1)
[25] Anderson B. 2011 There is no such thing as attention. Front. Psychol. 2, 246. (doi:10.3389/fpsyg.2011.00246)
[26] Corbetta M et al. 1998 A common network of functional areas for attention and eye movements. Neuron 21, 761–773. (doi:10.1016/S0896-6273(00)80593-0)
[27] Di Lollo V. 2018 Attention is a sterile concept; iterative reentry is a fertile substitute. Conscious Cogn. 64, 45–49. (doi:10.1016/j.concog.2018.02.005)
[28] Krauzlis RJ, Bollimunta A, Arcizet F, Wang L. 2014 Attention as an effect not a cause. Trends Cogn. Sci. 18, 457–464. (doi:10.1016/j.tics.2014.05.008)
[29] Tipper SP, Howard LA, Houghton G. 1998 Actionbased mechanisms of attention. Phil. Trans. R. Soc. Lond. B 353, 1385–1393. (doi:10.1098/rstb.1998.0292)
[30] Hommel B, Chapman CS, Cisek P, Neyedli HF, Song JH, Welsh TN. 2019 No one knows what attention is. Atten. Percept. Psychophys. 81, 2288–2303. (doi:10.3758/s13414-019-01846-w)
[31] Suryanarayana SM, Robertson B, Grillner S. 2021 The neural bases of vertebrate motor behaviour through the lens of evolution. Phil. Trans. R. Soc. B 377, 20200521. (doi:10.1098/rstb.2020.0521)
[32] Kozlov AK, Kardamakis AA, Hellgren Kotaleski J, Grillner S. 2014 Gating of steering signals through phasic modulation of reticulospinal neurons during locomotion. Proc. Natl Acad. Sci. USA 111, 3591–3596.(doi:10.1073/pnas.1401459111)
[33] Kardamakis AA, Saitoh K, Grillner S. 2015 Tecta microcircuit generating visual selection commands on gaze-controlling neurons. Proc. Natl Acad. Sci. USA 112, E1956–E1965. (doi:10.1073/pnas.1504866112)
[34] Saitoh K, Menard A, Grillner S. 2007 Tectal control of locomotion, steering, and eye movements in lamprey. J. Neurophysiol. 97, 3093–3108. (doi:10.1152/jn.00639.2006)
[35] Grillner S, Robertson B, Stephenson-Jones M. 2013 The evolutionary origin of the vertebrate basal ganglia and its role in action selection. J. Physiol. 591, 5425–5431. (doi:10.1113/jphysiol.2012.246660)
[36] Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. 2021 Olfaction in lamprey pallium revisited—dual projections of mitral and tufted cells. Cell Rep. 34, 108596. (doi:10.1016/j.celrep.2020.108596)
[37] Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. 2020 The evolutionary origin of visual and somatosensory representation in the vertebrate pallium. Nat. Ecol. Evol. 4, 639–651. (doi:10.1038/s41559-020-1137-2)
[38] Lacalli T. 2021 An evolutionary perspective on chordate brain organization and function: insights
from amphioxus, and the problem of sentience. Phil. Trans. R. Soc. B 377, 20200520. (doi:10.1098/rstb.2020.0520)
[39] Erwin DH, Laflamme M, Tweedt SM, Sperling EA, Pisani D, Peterson KJ. 2011 The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science 334, 1091–1097 (doi:10.1126/science.1206375)
[40] Bertrand S, Escriva H. 2011 Evolutionary crossroads in developmental biology: amphioxus. Development 138, 4819–4830. (doi:10.1242/dev.066720)
[41] Sattelle DB, Buckingham SD. 2006 Invertebrate studies and their ongoing contributions to neuroscience. Invert. Neurosci. 6,1–3. (doi:10.1007/s10158-005-0014-7)
[42] Yartsev MM. 2017 The emperor’s new wardrobe: rebalancing diversity of animal models in neuroscience research. Science 358, 466–469. (doi:10.1126/science.aan8865)
[43] Laurent G. 2020 On the value of model diversity in neuroscience. Nat. Rev. Neurosci. 21, 395–396.(doi:10.1038/s41583-020-0323-1)
[44] MacIver MA, Finlay BL. 2021 The neuroecology of the water-to-land transition and the evolution of
the vertebrate brain. Phil. Trans. R. Soc. B 377,20200523. (doi:10.1098/rstb.2020.0523)
[45] MacIver MA, Schmitz L, Mugan U, Murphey TD, Mobley CD. 2017 Massive increase in visual range preceded the origin of terrestrial vertebrates. Proc. Natl Acad. Sci. USA 114, E2375–E2384. (doi:10.1073/pnas.1615563114)
[46] Jacobs LF. 2021 How the evolution of air breathing shaped hippocampal function. Phil. Trans. R. Soc. B 377, 20200532. (doi:10.1098/rstb.2020.0532)
[47] Jacobs LF. 2012 From chemotaxis to the cognitive map: the function of olfaction. Proc. Natl Acad. Sci. USA 109, 10 693–10 700. (doi:10.1073/pnas.1201880109)
[48] Kaas J, Qi H-X, Stepniewska I. 2021 Escaping the nocturnal bottleneck, and the evolution of the dorsal and ventral streams of visual processing in primates. Phil. Trans. R. Soc. B 377, 20210293. (doi:10.1098/rstb.2021.0293)
[49] Cisek P. 2021 Evolution of behavioural control from chordates to primates. Phil. Trans. R. Soc. B 377, 20200522. (doi:10.1098/rstb.2020.0522)
[50] Pezzulo G, Parr T, Friston K. 2021 The evolution of brain architectures for predictive coding and active inference. Phil. Trans. R. Soc. B 377, 20200531. (doi:10.1098/rstb.2020.0531)
[51] Leopold DA, Averbeck BB. 2021 Self-tuition as an essential design feature of the brain. Phil. Trans. R. Soc. B 377, 20200530. (doi:10.1098/rstb.2020.0530)
[52] Wilson SP, Prescott TJ. 2021 Scaffolding layered control architectures through constraint closure: insights into brain evolution and development. Phil. Trans. R. Soc. B 377, 20200519. (doi:10.1098/rstb.2020.0519)
[53] Fine JM, Hayden BY. 2021 The whole prefrontal cortex is premotor cortex. Phil. Trans. R. Soc. B 377,(doi:10.1098/rstb.2020.0524)
[54] Hayden BY, Niv Y. 2021 The case against economic values in the orbitofrontal cortex (or anywhere else in the brain). Behav. Neurosci. 135, 192. (doi:10.1037/bne0000448)
[55] Barrett L, Henzi SP, Barton RA. 2021 Experts in action: why we need an embodied social brain hypothesis. Phil. Trans. R. Soc. B 377, 20200533. (doi:10.1098/rstb.2020.0533)
[56] Pessoa L, Medina L, Desfilis E. 2021 Refocusing neuroscience: moving away from mental categories and towards complex behaviours. Phil. Trans. R. Soc. B 377, 20200534. (doi:10.1098/rstb.2020.0534)
[57] Lindquist KA, Barrett LF. 2012 A functional architecture of the human brain: emerging insights from the science of emotion. Trends Cogn. Sci. 16, 533–540. (doi:10.1016/j.tics.2012.09.005)
[58] LeDoux JE. 2021 As soon as there was life, there was danger: the deep history of survival behaviours and the shallower history of consciousness. Phil. Trans. R. Soc. B 377, 20210292. (doi:10.1098/rstb.2021.0292)
[59] Bryer MAH et al. 2021 The evolution of quantitative sensitivity. Phil. Trans. R. Soc. B 377, 20200529. (doi:10.1098/rstb.2020.0529)
[60] Redish AD et al. 2021 Computational validity: using computation to translate behaviours across species. Phil. Trans. R. Soc. B 377, 20200525. (doi:10.1098/rstb.2020.0525)
